Какие снипы действуют в 2020 году: СНиП РФ 2015-2021 — Актуализированные действующие редакции строительных норм и правил
нормы строительства на дачном участке
Любое строительство проводится по определенным правилам. Неважно это многоэтажный дом или веранда.Какие нормы строительства на дачном участке? Какие акты нужно оформить? Какие разрешения получить? Как возвести дачу, чтобы она соответствовала современным экологическим нормам? Как не нарушить территорию соседних участков?
Вопросов которые сопровождают строительство очень много. Попробуем разобраться на примере возведения здания на дачном участке.
Закон
Нормы строительства на дачном участке — это требования предъявляемые к возведению любого строения. При покупке земли у владельца самые широкие полномочия по ее использованию. Но СНиПы и Госты по строительству действующие в 2021 году вносят свои коррективы. Если не соблюдать правила то собственника ждут большие штрафы, вплоть до снесения построек.
Какие нормы стоит соблюдать при строительстве дачного дома? Полагаться следует на Градостроительные кодекс и СНиП 30-02-97 в действующей редакции.
Для справки можно обратиться дополнительно к ФЗ №66 «О садоводческих, огороднических и дачных некоммерческих объединений граждан».
ВНИМАНИЕ! Нормы указанные в СНиП 30-02-97 от 2001 года обязательны к исполнению, а требования в документе СП 11-106-97 представлены для ознакомления и соблюдаются по желанию.
В настоящее время Налоговая служба ужесточила контроль за частным строительством.
Но, как правило, на участке несколько построек. Какие из них можно считать капитальными?
- Дом, в котором проживает семья.

- Гараж с фундаментом.
- Баня.

Разрешения не требуются при строительстве веранды, беседки, навеса и т.д. Что касается террасы. Если она открытая или прилегает к дому, то дополнительных согласований не требуется. Но когда возведение подразумевает обнесение стеной с крышей, то без разрешения не обойтись.
Перечислим основной пакет документов для разрешения под ИЖС:
- Документ, удостоверяющий личность.
- Заявление на регистрацию.
- Свидетельство о праве собственности.
- Технический и межевой планы участка.
- Кадастровый паспорт.
- Справку об отсутствии обременений.
- Разрешение от органов местного самоуправления и прочих служб.
Нормы строительства на дачном участке
Расположение садовых товариществ на карте выглядит, как правило, компактно. Загородные дома часто не предполагают постоянного проживания.
 Дачные постройки небольшие с минимум удобств, чтобы отдохнуть после тяжелых огородных дел.
Дачные постройки небольшие с минимум удобств, чтобы отдохнуть после тяжелых огородных дел.ВНИМАНИЕ! Постановления и нормы о дачных строения приходится учитывать задолго до начала строительства. В идеале на стадии планирования и оформления бумаг.
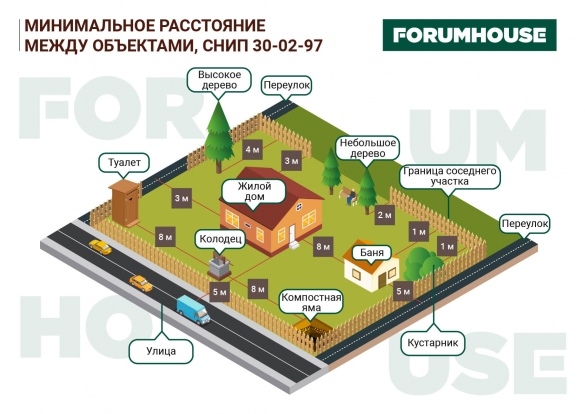
Строительные правила указанные в СНиП четко регулируют положение загородного дома на участке, забора и расстояния между объектами. Обратимся к основным нормам строительства:
- Минимальная высота для загородной дачи — 2,2 м. Для жилого дома — 2,5 м.
- При временной недоступности канализации предусмотрено отведение места под компостную яму. Она располагается не ближе чем на 8 м к колодцу или входу в дом. Но, как правило, ее организовывают вблизи сельского туалета. Расстояние от дачной постройки до бани или душа под открытым небом составляет 8 м.
- В обязательном порядке для возведения требуется источник пресной воды. Как ни странно, свет и газ вовсе не на первых ролях. Причем, воду можно брать из уличной колонки или родника (реки, озера и т.д.).
- Взаимоотношения с соседними строениями крайне важны. Участок огораживается забором или сеткой в обязательном порядке. Нельзя просто так поставить высокое заграждение, чтобы обезопасить себя от посторонних глаз.
 Причем, воду можно брать из уличной колонки или родника (реки, озера и т.д.).
Причем, воду можно брать из уличной колонки или родника (реки, озера и т.д.).Высокий, глухой забор допускается со стороны дороги или по согласованию с соседями и председателем садового товарищества. По правилам ограждение не должно быть выше, чем 1,5 м. Например, расстояние от забора до сарая с крупным или мелким рогатым скотом составляет не менее 12 метров.
Очень важно чтобы при отсутствии канализации все стоки отводились в специальную траншею из мелкого песка или гравия.
Правил немало и все они обязаны к выполнению. Но как быть если дачные постройки расположены близко к друг другу, практически вплотную?  Рассмотрим на примере гаража:
Рассмотрим на примере гаража:
- Постройка для автомобиля обязательно имеет отдельный вход. Даже если в гараж можно попасть через дверь в стене дома.
- Расстояние от забора до одного края дома — до 3-х метров, до гаражной пристройки — до 1 метра.
- Вход в автомобильную постройку должен быть не меньше 7 метров от входа в дом.
СНиП 30-02-97 от 2001 года так же регулирует расстояние между дачными постройками в целях пожарной безопасности.
- Расстояние между бетонными или кирпичными строениями — 6 метров.
- Расстояние между кирпичными и деревянными строениями — 8 метров и более.
- Расстояние между деревянными строениями — 15 метров и более.
Законом предусмотрено, что любой садовый дом должен соответствовать определенному расстоянию от заборами и между самими постройками:
- От основного здания до соседнего участка —
- От бани, сарая или гаража до соседского забора — 1 метр.
- От сарая предназначенного для мелкого скота (загона) — 4 метра.

Многие дачники любят окружать себя зелеными насаждениями. Этот момент так же регулируется т.к взаимоотношения с соседними строениями один из главных пунктов правил:
- Расстояние до границ соседнего участка должно быть не менее 3 метров, а до другой постройки не менее 4 метров. В зависимости от высоты дерева можно сократить посадку до 2 метров.
- Кустарники следует выращивать не ближе 1 метра от границ соседней земли.
Нюансы расположения построек
Некоторые основные нормы мы указали выше. Застройка дачного участка содержит ряд минусов. Главный из них — площадь. Часто очень сложно расположить строения на ограниченной территории. Но нарушение норм приведет к серьезным последствиям.
ВАЖНО! Минимальный размер земельного участка пригодного для строительства — 6 соток.

Практика показала, что лучшая форма для дачного участка квадратная. Она максимально подходит для компактного и комфортного размещения строений даже при дефиците площади.
С другой стороны покупка большого участка земли выйдет недешево. Причем, в перспективе. Поддержание и обслуживание огромного особняка дорогое удовольствие. На участке в 6 соток максимально допустимые размеры дачного домика будут около 130 м2.
Как быть с постройками, которые предусматривают выступающую крышу или навес? Расстояние до 0,5 метра обычно не принимают по внимание. Такой выступ не создает неудобств.Для отчета можно выбрать стену или цоколь при его наличии.
 Соответственно забор стоит устанавливать с учетом дополнительной величины.
Соответственно забор стоит устанавливать с учетом дополнительной величины.Парковочное место для автомобиля занимает, как правило, до 1/4 от общей площади. Сюда же входят дорожки. Хотя документы регулируют плотность застройки для участка в 6 соток не более 30%.
«Красная линия» — граница которая отделяет общественные постройки от дачных. Если дом стоит на полноценной улице, то расстояние не должно превышать 5 метров. Минимальная граница очерчена в 3 метра.
Обратим внимание на действующий в 2021 году СНиП, который регулирует инженерное оборудование:
- На территории участка запрещено размещать баллоны с газом. В доме при отсутствии центрального обеспечения разрешается в единственном экземпляре объемом не более 12 л.
- В основной постройке в обязательном порядке устанавливается счетчик электроэнергии.
- Нормы предполагают наличие стока, чтобы избежать попадания дождевой воды на соседний участок.
 В доме при отсутствии центрального обеспечения разрешается в единственном экземпляре объемом не более 12 л.
В доме при отсутствии центрального обеспечения разрешается в единственном экземпляре объемом не более 12 л.Что касается погреба или подвала. Он запрещен для строительство под баней или сараем. Он должен располагаться под дачным домом. Высота подвала — не менее 2,2 м.
Нормы строительства обязывают граждан иметь на дачном участке радио. Делается это для своевременного оповещения во время чрезвычайных ситуаций.
Действия при нарушении
К сожалению, не все граждане исполняют строгие правила. Нарушения часто становятся причиной пожаров или приносят материальный ущерб соседям.
ВНИМАНИЕ! Простое отсутствие дождевого стока может привести к гибели урожая или затоплению дома.
Что делать, если соседний участок нарушает нормы строительства? Например, туалет расположен ближе чем 3 метра от колодца:
- С помощью фото- или видеосъемки зафиксировать нарушение.
- Составить акт и обратиться в СНТ с доказательствами. СНТ выделит гражданину время для устранения нарушений.
- Если гражданин проигнорировал законные требования, а срок на исправление вышел и нет адекватных причин для его продления, то бумаги передают в вышестоящие органы. Возможно содействие с пожарной службой и архитектурной инспекцией.
- В крайнем случае следует обращение в суд. Подается иск с приложенным актом о нарушении, доказательствами, заключением экспертов.

Соблюдение норм строительства на дачном участке позволит обезопасить себя от чрезвычайных происшествий, сделать свою жизнь комфортней и избежать конфликтных ситуаций с соседями и надзорными органами. Грамотное и своевременное планирование сэкономит деньги и время.
Какие СП пришли на замену СНиПам о строительстве дома — Реальное время
Где найти нормы и правила, по которым должно идти проектирование и строительство
Главные параметры того, что же в Российской Федерации вообще может называться жилым домом (а значит, может быть зарегистрировано в его качестве), обрисовываются в своде правил «Дома жилые одноквартирные». В нем даются ссылки и на санитарно-гигиенические требования, и на технические регламенты о требованиях пожарной безопасности, и на многочисленные другие своды правил, регламентирующие разные аспекты индивидуального дома, его строительства и жизни в нем. Словом, именно от этого СП нужно «танцевать», определяя основные контуры вашего будущего дома.
В нем даются ссылки и на санитарно-гигиенические требования, и на технические регламенты о требованиях пожарной безопасности, и на многочисленные другие своды правил, регламентирующие разные аспекты индивидуального дома, его строительства и жизни в нем. Словом, именно от этого СП нужно «танцевать», определяя основные контуры вашего будущего дома.
За общую отрисовку нормативов по жилому дому в России отвечает свод правил «Дома жилые одноквартирные» — СП 55.13330.2016, который действует с 21 апреля 2017 года. Этот документ революцией не стал — он лишь является актуализированной редакцией СНиП 31-02-2001, действовавшего прежде.
Для начала в своде правил объясняется, из чего вообще должен состоять дом: в нем должны быть жилые комнаты, кухня, ванная комната (или душевая), туалет, кладовая или встроенные шкафы. А если участок не включается в систему централизованного теплоснабжения — нужно отдельное помещение. В доме для постоянного проживания нужно обязательно предусмотреть отопление (с температурой не ниже 20 градусов для всех помещений с постоянным пребыванием людей), вентиляцию, водоснабжение, канализацию и электроснабжение.
В доме для постоянного проживания нужно обязательно предусмотреть отопление (с температурой не ниже 20 градусов для всех помещений с постоянным пребыванием людей), вентиляцию, водоснабжение, канализацию и электроснабжение.
СП 55.13330.2016 устанавливает и минимальные габариты, которым должен соответствовать жилой дом. Вот что рекомендуется в этом документе:
- Площадь общей комнаты — 12 кв. м.
- Спальня должна быть не меньше 8 кв. м (но если она находится в мансарде — допускается обойтись и 7 кв. м).
- Минимальная площадь кухни — 6 кв. м.
- Ширина помещений тоже регламентируется: кухня — не уже 1,7 метра, коридор — не уже 1,4 метра. На ширину ванной комнаты отводится не менее полутора метров, а туалету хватит и 0,8 м. А вот глубина туалета должна быть не менее 1,2 м (если дверь открывается наружу) или 1,5 м (если дверь открывается внутрь).
Высота потолка в жилых комнатах и кухнях зависит от того, к какому климатическому району относится регион в соответствии со СНиП 23-01-99 («Строительная климатология»). В Татарстане эта цифра должна быть не менее 2,5 метра. В коридорах высоту можно уменьшать до 2,1 метра, а в мансарде — лишь до 2,3 метра.
В Татарстане эта цифра должна быть не менее 2,5 метра. В коридорах высоту можно уменьшать до 2,1 метра, а в мансарде — лишь до 2,3 метра.
А вот что касается максимальной площади дома — то она теоретически не ограничивается. Правда, если его площадь более 500 квадратных метров, то уведомительный характер строительства действовать уже не будет. В этом случае нужно будет разрабатывать и согласовывать проектную документацию, проводить ее через экспертизу и получать разрешение на ввод в эксплуатацию (а это гораздо сложнее, чем уведомительная процедура, которую мы описывали в предыдущих статьях).
Фундаментальный вопросОдин из ключевых параметров дома — фундамент и распределение нагрузок. Вряд ли вы будете рассчитывать все эти параметры самостоятельно, ведь этим занимаются специально обученные люди (архитекторы-проектировщики).
Однако полезно будет знать, на что они опираются в своих расчетах. Есть СП 20.13330.2011, в котором детально прописываются расчеты нагрузок и воздействий. В этом своде правил учтен каждый вид нагрузки на конструкции зданий и сооружений, начиная от наличия в здании людей и заканчивая ветровой нагрузкой. Этот свод правил изобилует формулами для расчетов. Если обычному человеку они кажутся китайской грамотой, то архитектор, разрабатывая проект дома, детально выяснит, какая нагрузка приходится на конкретную стену или, скажем, колонну, как фундамент будет реагировать на тот или иной тип перекрытий, какая снеговая нагрузка ляжет на вашу крышу и т.д. В этом же СП рассчитываются температурные и климатические воздействия, приводятся разнообразные таблицы (например, как поглощают солнечную радиацию наружные поверхности из разных материалов). Все эти расчеты впоследствии лягут в основу проекта дома.
В этом своде правил учтен каждый вид нагрузки на конструкции зданий и сооружений, начиная от наличия в здании людей и заканчивая ветровой нагрузкой. Этот свод правил изобилует формулами для расчетов. Если обычному человеку они кажутся китайской грамотой, то архитектор, разрабатывая проект дома, детально выяснит, какая нагрузка приходится на конкретную стену или, скажем, колонну, как фундамент будет реагировать на тот или иной тип перекрытий, какая снеговая нагрузка ляжет на вашу крышу и т.д. В этом же СП рассчитываются температурные и климатические воздействия, приводятся разнообразные таблицы (например, как поглощают солнечную радиацию наружные поверхности из разных материалов). Все эти расчеты впоследствии лягут в основу проекта дома.
Еще один свод правил — один из важнейших, на который ориентируются строители, — СП 22.13330.2016 «Основания зданий и сооружений». Именно по этим правилам рассчитываются фундаменты зданий, включая и наши индивидуальные дома. Это весьма подробный документ, разбираться в котором должен специалист по строительству и проектированию. Здесь приводятся формулы расчетов фундамента (их требуемой прочности, сопротивления, предельных нагрузок, возможности деформации и т.д.) в зависимости от разных факторов (основной из которых — особенности грунта). В нашем основном СП сказано, что именно на СП 22.13330.2016 нужно ориентироваться при проектировании фундамента дома, в зависимости от того, какой у нас на участке грунт и гидрогеологический режим.
Здесь приводятся формулы расчетов фундамента (их требуемой прочности, сопротивления, предельных нагрузок, возможности деформации и т.д.) в зависимости от разных факторов (основной из которых — особенности грунта). В нашем основном СП сказано, что именно на СП 22.13330.2016 нужно ориентироваться при проектировании фундамента дома, в зависимости от того, какой у нас на участке грунт и гидрогеологический режим.
Согласно действующему Техническому регламенту о требованиях пожарной безопасности, к одноэтажным и двухэтажным домам не предъявляется никаких требований по степени огнестойкости и классу конструктивной пожарной опасности. А вот если этажей в нашем доме 3, то стоит заглянуть в таблицу 21 этого самого регламента: там перечислены требования к огнестойкости несущих элементов (не менее R 45), перекрытий (REI 45), ненесущих наружных стен (E15) и прочих элементов. Не регламентируется в таком доме только огнестойкость межкомнатных перегородок.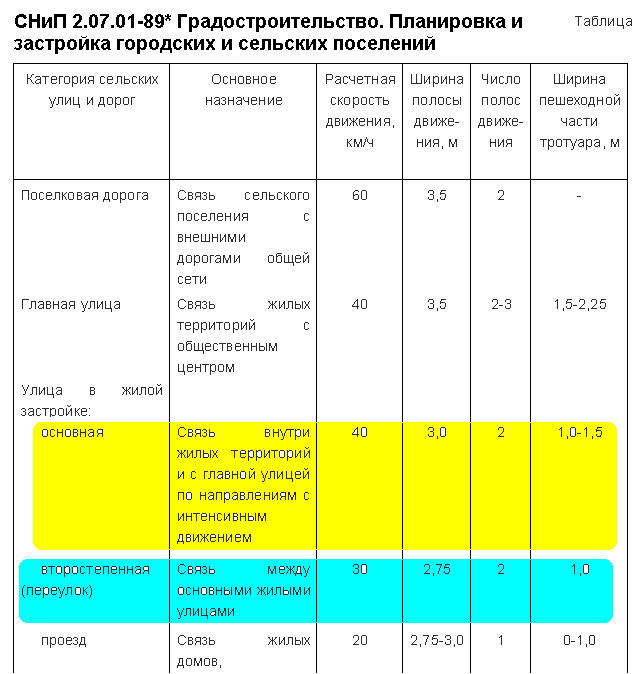 Общий класс конструктивной пожарной опасности трехэтажного дома должен быть не ниже С2.
Общий класс конструктивной пожарной опасности трехэтажного дома должен быть не ниже С2.
А еще по этому регламенту каждый дом должен иметь минимум один эвакуационный выход на улицу. Трехэтажный дом должен быть оборудован автономными дымовыми пожарными извещателями, причем на каждом этаже (правда, их не следует ставить на кухне, в ванных комнатах, душевых и туалетах).
Кроме прочего, в основном СП, посвященном строительству индивидуальных домов, подробно прописывается, как должны быть организованы камины, печи, варочные плиты, дымоходы (пп. 6.13 и 6.14).
Общая безопасность дома и системы комфортаДом, согласно своду правил, в принципе должен быть безопасным сооружением. Поэтому заботливые составители Свода правил отдельно прописали, как нужно об этом позаботиться. Из интересного и полезного:
- Не допускается применение лестниц с разной высотой ступеней;
- Высота ограждений лестниц, балконов, лоджий, кровли должна быть не ниже 90 см;
- Температура поверхностей доступных частей нагревательных приборов не должна быть выше 70 градусов по Цельсию, если до них можно дотронуться напрямую, и выше 90 градусов, если они забраны защитным коробом. Если отопление воздушное, то горячий воздух, выходящий из выдувного отверстия, ограничивается температурой в 70 градусов.
- Температура горячей воды в ваших кранах не должна быть выше 60 градусов.
 Если отопление воздушное, то горячий воздух, выходящий из выдувного отверстия, ограничивается температурой в 70 градусов.
Если отопление воздушное, то горячий воздух, выходящий из выдувного отверстия, ограничивается температурой в 70 градусов.
Достаточно вскользь упоминаются технические системы дома: как должна быть организована система вентиляции (например, из кухни в режиме обслуживания должно удаляться не менее 60 кубометров воздуха в час, а из ванной и туалета — 25 кубометров), отопление (СНиП 41-01-2003), звукоизоляция (здесь документ ссылается на СП 51.13330.2011 «Защита от шума»).
Освещение дома — важная часть его проекта. Здесь нужно ориентироваться на СП 52.13330.2011 «Естественное и искусственное освещение» и на СанПиН 2.1.2.2645-10 (раздел 5). В жилых комнатах и на кухне должно быть обеспечено естественное освещение (через окна). Определено сводом правил даже отношение площади световых проемов к площади пола в таких помещениях, и оно должно быть не меньше 1:8 (п. 8.7).
Словом, в своде правил, посвященном индивидуальному жилому дому, много интересной информации, но в основном идут отсылки к другим действующим СП, СанПиН и ГОСТам. Каждый отдельный вопрос по строительству описан в собственном регламентирующем документе, и в дальнейшем мы обязательно будем более подробно на них ссылаться и их изучать.
Каждый отдельный вопрос по строительству описан в собственном регламентирующем документе, и в дальнейшем мы обязательно будем более подробно на них ссылаться и их изучать.
Людмила Губаева
Недвижимость ТатарстанВЗГЛЯД / Каким должно быть расстояние между объектами на даче по новым правилам? :: Вопрос дня
В середине апреля в России вступил в силу новый свод правил (СП), касающихся застройки садовых участков. Из-за пандемии это событие прошло почти незамеченным, и отчасти незаслуженно, поскольку немалое количество россиян проектирует свои участки с оглядкой на СП.
С 1998 года в России действовал СНиП 30-02-97, который регламентировал все дачное строительство: какие дома строить, на каком расстоянии от забора, какой эти самые заборы должны быть высоты, где расположить баню, компостные ямы и туалеты, а также многое другое. Его несколько раз актуализировали, последняя редакция была в 2011 году.
С 15 апреля 2020-го в стране действует новая версия – СП 53.13330.2019, в которой многие нормы пересмотрели.
Основных изменения, пожалуй, два. Первое касается минимально допустимого размера участка. Было шесть соток (0,06 га), стало четыре (0,04). Это не значит, что всем дачникам придется уменьшать свои участки – новые правила обратной силы не имеют, так что касаются они лишь планировки новых СНТ и построек, которые пока не возведены.
Но по большому счету это изменение – лишь формальность, поскольку в России минимально и максимально допустимые размеры земельных участков, в том числе предназначенных для садоводства, устанавливают власти регионов. В Вологодской области, к примеру, вполне законны участки даже в три сотки.
То есть участки меньше шести соток в стране давно являются нормой, и Минстрой своим новым сводом правил это просто зафиксировал. А это изменение, в свою очередь, потребовало и пересмотра допускавшихся ранее расстояний между объектами, часть из них сократили.
Исключение касается расположения септиков, отдельно стоящих туалетов и компостных ям по отношению к соседскому участку. Раньше этот момент вообще никак не оговаривался – в СП было лишь прописано, что между постройками вроде туалета на вашем участке и соседским забором должно было быть не менее метра. Сейчас же в своде правил отдельно упомянули этот момент – от соседского забора до туалета, септика или компостной кучи должно быть как минимум два метра. А до стен соседского дома – и вовсе 12 метров. Это второе важное изменение.
Остальные – более мелкие. Например, крупные высокие деревья теперь могут быть на метр ближе к чужому забору – в трех метрах максимум. Расстояния внутри участка остались прежними – от дома до бани, душа, туалета или компоста – минимум восемь метров, от колодца до туалета или компоста – тоже восемь. До границы с соседями от дома должно быть три метра, от крупных хозпостроек вроде курятников или сараев – четыре, от кустарников – один метр.
Изменилась максимально допустимая высота заборов: была полтора метра, теперь – 1,8. Причем в документе отдельно прописано, что заборы по всему периметру участка рекомендуется делать сетчатыми, а не глухими. Иной вариант допускается только при письменном согласии соседей.
Причем в документе отдельно прописано, что заборы по всему периметру участка рекомендуется делать сетчатыми, а не глухими. Иной вариант допускается только при письменном согласии соседей.
И этот момент довольно показателен. Любой россиянин знает, что далеко не все заборы в стране сетчатые. Скорее можно сказать, что их минимум. И дело здесь не только в том, что большую их часть возвели еще до появления нынешних СНиПов и СП, многие появились и совсем недавно. Дело в том, что ни актуальный свод правил по застройке дачных участков, ни его предшественники не являются документами, обязательными к исполнению.
В России есть федеральный закон «Требования к безопасности зданий и сооружений», к которому прилагается перечень под номером 1521 из более чем 70 ГОСТов и сводов правил, бывших СНиПов. Вот этим стандартам и правилам следовать нужно обязательно. СП 53.13330.2019 в их числе нет.
Он, точнее его предшественник, есть в другом списке, утвержденном приказом Росстандарта от апреля 2019 года. Это перечень ГОСТов и СП, которые можно применять добровольно.
Это перечень ГОСТов и СП, которые можно применять добровольно.
Минстрой даже сам косвенно подтверждал, что правилам застройки дач следовать совсем не обязательно. В 2015 году в письме-ответе на один из запросов говорилось, что все правила и стандарты, не включенные в список 1521, «утрачивают обязательность применения и применяются на добровольной основе».
Так что следовать СП 53.13330.2019 хоть и стоит, ведь он составлен в том числе для того, чтобы сделать жизнь в садовых товариществах более комфортной, но делать это совсем не обязательно. И более того, решить спор в суде этот свод тоже не поможет, сослаться на него не получится как раз в силу его «добровольности». Единственный вариант – доказать, что, условно, нечистоты из стоящего вплотную к вашему забору соседского туалета отравили воду в вашем колодце. То есть показать суду конкретный ущерб.
Новые 14 строительных норм (СНиП) приняты в Беларуси в 2020 году
Постановление Министерства строительства и архитектуры РБ от 16 декабря 2019 года № 69 вступит в силу с 8 сентября 2020 года
Документом утверждается вступление в силу 14 новых строительных норм (СНиП) по следующим разделам:
- Жилые здания;
- Общественные здания;
- Кровли;
- Станции технического обслуживания транспортных средств. Гаражи-стоянки автомобилей;
- Воздействия на конструкции. Общие воздействия. Воздействия для определения огнестойкости;
- Воздействия на конструкции. Общие воздействия. Ветровые воздействия;
- Воздействия на конструкции. Общие воздействия. Объемный вес, собственный вес, функциональные нагрузки для зданий
- Воздействия на конструкции. Общие воздействия. Температурные воздействия;
- Воздействия на конструкции. Общие воздействия. Снеговые нагрузки;
- Отопление, вентиляция и кондиционирование воздуха;
- Основы проектирования строительных конструкций;
- Промышленный транспорт;
- Трамвайные и троллейбусные линии;
- Склады нефти и нефтепродуктов.
 Гаражи-стоянки автомобилей;
Гаражи-стоянки автомобилей;Данные изменения в строительное законодательство разработаны Республиканским унитарным предприятием «Стройтехнорм» и согласованы с главным управлением градостроительства, проектной, научно-технической и инновационной политики Министерства строительства и архитектуры Беларуси.
В Белоруссии идет масштабное изменение строительного законодательства
Согласно Указа Президента Республики Беларусь от 5 июня 2019 года № 217. Строительные нормы. в целях упорядочения требований технических нормативных правовых актов в сфере архитектуры, градостроительства и строительства устанавливается, что обязательные для соблюдения требования в области архитектурной, градостроительной и строительной деятельности при градостроительном планировании, размещении объектов строительства и застройке территорий, проектировании и строительстве зданий и сооружений устанавливаются в строительных нормах в целях обеспечения:
- механической прочности и устойчивости зданий и сооружений;
- пожарной, промышленной, ядерной, радиационной, энергетической безопасности;
- защиты от чрезвычайных ситуаций природного и техногенного характера;
- экономии энергии и тепловой защиты;
- нормативов охраны окружающей среды и санитарно-эпидемиологического благополучия населения;
Текст указа Президента РБ №217 от 05.
 06.2019 г. прочтите ниже.
06.2019 г. прочтите ниже.Действующие, на данный момент, технические нормативные правовые акты других государственных органов в части, устанавливающей обязательные требования при проектировании и строительстве объектов, будут признаваться утратившими силу после введения в действие новых строительных норм и правил (СНиП).
Новые СНиПы начнут действовать с сентября 2020 года
Обновленные СНиП по 14 разделам строительных норм и правил вступят в силу через 60 дней после опубликования постановления Минстройархитектуры №69 от 16.12.2019 года на Национальном правовом интернет-портале Республики Беларусь (Опубликованы сегодня 09.07.2020 г.).
Ранее, в январе 2020 года Постановлением № 24/33 Министерства труда и социальной защиты и Минстройархитектуры РБ от 31.05.2019 вступили в силу новые Правила по охране труда при выполнении строительных работ в Республике Беларусь.
Поделиться ссылкой:
СНиПы, ГОСТы, СП – как разобраться и чем руководствоваться при проектировании «Доступной среды»?
У нас часто спрашивают, какими нормами нужно руководствоваться, чтобы оборудовать объект по программе «Доступная среда».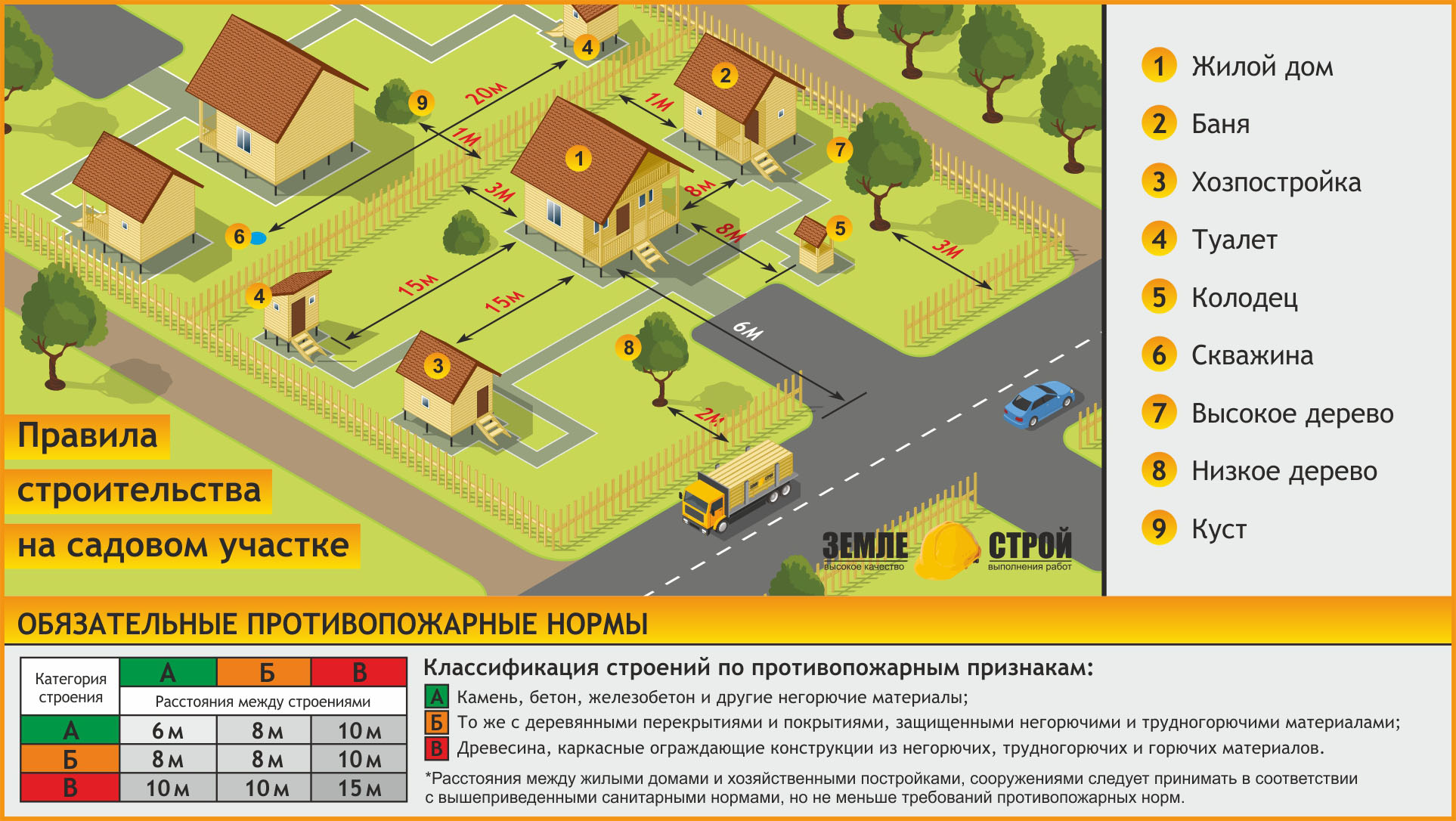 Где уточнить правильный угол наклона пандуса, высоту расположения тех или иных знаков, нормативы установки поручней? Где найти актуальные редакции всех документов, которые часто устаревают и обновляются далеко не на всех ресурсах?
Где уточнить правильный угол наклона пандуса, высоту расположения тех или иных знаков, нормативы установки поручней? Где найти актуальные редакции всех документов, которые часто устаревают и обновляются далеко не на всех ресурсах?
Самый правильный вариант – доверить работу с технической документацией проектировщикам компании «Неограниченные возможности». Наши специалисты прошли специальное обучение и обладают необходимыми навыками для подготовки проекта любой сложности.
Специально для вас мы собрали ссылки на все действующие нормативы по государственной программе «Доступная среда». Именно их используют наши специалисты в своей работе.
Документ о правах инвалидов для общего ознакомления:
Конвенция о правах инвалидов принята резолюцией 61/106 Генеральной Ассамблеи ООН от 13 декабря 2006 года, вступила в силу 3 мая 2008 года.
Основной документ, касающийся строительства – это свод правил СП 59.13330.2016 Доступность зданий и сооружений для маломобильных групп населения.
Руководящие документы системы РДС 35-201-99 Порядок реализации требований доступности для инвалидов к объектам социальной инфраструктуры.
Ведомственные строительные нормы ВСН 62-91 Проектирование среды жизнедеятельности с учетом потребностей инвалидов и маломобильных групп населения.
СВОДЫ ПРАВИЛ
СП 31-102-99 Требования доступности общественных зданий и сооружений для инвалидов и других маломобильных посетителей.
СП 35-101-2001 Проектирование зданий и сооружений с учетом доступности для маломобильных групп населения. Общие положения.
СП 35-102-2001 Жилая среда с планировочными элементами, доступными инвалидам.
СП 35-103-2001 Общественные здания и сооружения, доступные маломобильным посетителям.
СП 35-104-2001 Здания и помещения с местами труда для инвалидов.
СП 35-105-2002 Реконструкция городской застройки с учетом доступности для инвалидов и других маломобильных групп населения.
СП 35-106-2003 Расчет и размещение учреждений социального обслуживания пожилых людей.
СП 35-114-2003 Свод правил по проектированию и строительству. Реконструкция и приспособление зданий для учреждений социального обслуживания пожилых людей.
СП 35-115-2004 Свод правил по проектированию и строительству. Обустройство помещений в учреждениях социального и медицинского обслуживания пожилых людей.
Новые ГОСТы, которые вводятся в 2020 году
Мы следим за новыми Гостами, связанными с нашими видами работ и вообще с промышленной безопасностью, и обновляем на этой странице:
Неразрушающий контроль
| ==================== | |||
| ГОСТ Р 56542-2019 | Контроль неразрушающий. Классификация видов и методов | 01.11.2020 | |
| ГОСТ Р 58713-2019 | Контроль неразрушающий. Несплошности в образцах, используемых в квалификационных экзаменах | 01.03.2020 | |
| ГОСТ Р 58764-2019 | Контроль неразрушающий. Методы оптические. Эндоскопы технические. Общие требования | 01.03.2020 | |
| ГОСТ Р ИСО 9712-2019 | Контроль неразрушающий. Квалификация и сертификация персонала | 01.03.2020 | |
| ГОСТ Р 8.969-2019 | Государственная система обеспечения единства измерений. Металлы и сплавы. Определение твердости по шкалам Либа. Часть 1. Метод измерений | 01.03.2020 | |
Надежность, оценка риска
| ==================== | |||
| ГОСТ Р 27.012-2019 | Надежность в технике. Анализ опасности и работоспособности (HAZOP) | 01.07.2020 | |
| ГОСТ Р 27.013-2019 | Надежность в технике. Методы оценки показателей безотказности | 01.07.2020 | |
| ГОСТ Р 27.014-2019 | Надежность в технике. Управление надежностью. Руководство по установлению требований к надежности систем | 01.07.2020 | |
| ГОСТ Р 27.015-2019 | Надежность в технике. Управление надежностью. Руководство по проектированию надежности систем | 01.07.2020 | |
ГОСТ Р 58771-2019 | Менеджмент риска. Технологии оценки риска | 01.03.2020 |
Здания и сооружения
| ==================== | ||
| ГОСТ Р 22.1.03-2019 | Национальный стандарт Российской Федерации. Безопасность в чрезвычайных ситуациях. Система мониторинга инженерных систем зданий и сооружений. Технические требования. Протоколы информационного обмена | 01.04.2020 |
Подъемные сооружения
| ==================== | |||
| ГОСТ 34466-2018 | Краны грузоподъемные. Требования к компетентности крановщиков (операторов), стропальщиков и сигнальщиков | 01.06.2020 | |
| ГОСТ 34465.1-2018 | Краны грузоподъемные. Органы управления. Расположение и характеристики. Часть 1. Общие положения | 01.06.2020 | |
| ГОСТ 34465.2-2018 | Краны грузоподъемные. Органы управления. Расположение и характеристики. Часть 2. Краны стреловые самоходные | 01.06.2020 | |
| ГОСТ 34465.3-2019 | Краны грузоподъемные. Органы управления. Расположение и характеристики. Часть 3. Краны башенные | 01.06.2020 | |
| ГОСТ 34465.4-2018 | Краны грузоподъемные. Органы управления. Расположение и характеристики. Часть 4. Краны стреловые | 01.06.2020 | |
| ГОСТ 34585-2019 | Краны грузоподъемные. Обучение стропальщиков и сигнальщиков | 01.06.2020 | |
| ГОСТ 31272.1-2018 | Краны грузоподъемные. Обучение крановщиков (операторов). Часть 1. Общие положения | 01.06.2020 | |
| ГОСТ 31272.3-2019 | Краны грузоподъемные. Обучение крановщиков (операторов). Часть 3. Краны башенные | 01.06.2020 | |
| ГОСТ 34463.1-2018 | Краны грузоподъемные. Безопасная эксплуатация. Часть 1. Общие положения | 01.06.2020 | |
| ГОСТ 34463.3-2019 | Краны грузоподъемные. Безопасная эксплуатация. Часть 3. Краны башенные | 01.06.2020 | |
| ГОСТ 34463.4-2018 | Краны грузоподъемные. Безопасная эксплуатация. Часть 4. Краны стреловые | 01.06.2020 | |
| ГОСТ 34464.1-2018 | Краны грузоподъемные. Информация, предоставляемая по запросу. Часть 1. Общие положения | 01.06.2020 | |
| ГОСТ 34464.3-2019 | Краны грузоподъемные. Информация, предоставляемая по запросу. Часть 3. Краны башенные | 01.06.2020 | |
| ГОСТ 34464.4-2018 | Краны грузоподъемные. Информация, предоставляемая по запросу. Часть 4. Краны стреловые | 01.06.2020 | |
| ГОСТ 34586.1-2019 | Краны грузоподъемные. Графические символы. Часть 1. Общие положения | 01.06.2020 | |
| ГОСТ 34586.2-2019 | Краны грузоподъемные. Графические символы. Часть 2. Краны стреловые самоходные | 01.06.2020 | |
| ГОСТ 34586.3-2019 | Краны грузоподъемные. Графические символы. Часть 3. Краны башенные | 01.06.2020 | |
| ГОСТ 34584-2019 | Краны грузоподъемные. Измерение массы крана и его компонентов | 01.06.2020 | |
| ГОСТ 34587-2019 | Краны грузоподъемные. Металлические конструкции. Требования к изготовлению | 01.06.2020 | |
| ГОСТ 34588-2019 | Краны грузоподъемные. Предупреждающие знаки и пиктограммы. Общие принципы | 01.06.2020 | |
| ГОСТ 34589-2019 | Краны грузоподъемные. Краны мостовые и козловые. Общие технические требования | 01.06.2020 | |
| ГОСТ 34591-2019 | Краны грузоподъемные. Ручные сигналы | 01.06.2020 | |
| ГОСТ Р 58520-2019 | Средства грузозахватные. Классификация и общие технические требования | 01.03.2020 | |
| ГОСТ Р 58753-2019 | Национальный стандарт Российской Федерации. Стропы грузовые канатные для строительства. Технические условия | 01.09.2020 |
Газ, котлы
| ==================== | |||
| ГОСТ 34608-2019 | Устройства безопасности и управления для газовых горелок и приборов, сжигающих газ. Клапаны запорные автоматические для рабочего давления свыше 500 до 6300 кПа включительно | 01.05.2020 | |
| ГОСТ 34609-2019 | Краны шаровые и краны с конусным закрывающим элементом с ручным управлением для газовых установок зданий | 01.05.2020 | |
| ГОСТ Р 53865-2019 | Системы газораспределительные. Термины и определения | 01.05.2020 | |
| ГОСТ Р 55474-2019 | Системы газораспределительные. Сети газораспределения природного газа. Часть 2. Стальные газопроводы» | 01.06.2020 | |
| ГОСТ Р 58778-2019 | Системы газораспределительные. Сети газораспределения и газопотребления. Газопроводы высокого давления категории 1а | 01.05.2020 | |
| ГОСТ 20548-93 | Котлы отопительные водогрейные теплопроизводительностью до 100 кВт. Общие технические условия | 01.01.2021 |
Лифты
| ==================== | |||
| ГОСТ 8824-2018 | Лифты грузовые малые. Основные параметры и размеры | 01.06.2020 | |
| ГОСТ Р 58495-2019 | Лифты. Модернизация находящихся в эксплуатации лифтов. Общие требования | 01.01.2020 | |
| ГОСТ 34486.1-2018 | Лифты. Правила и методы исследований (испытаний) и измерений. Измерение качества движения | 01.06.2020 | |
| ГОСТ 34486.2-2018 | Эскалаторы и пассажирские конвейеры. Правила и методы исследований (испытаний) и измерений. Измерение качества движения | 01.06.2020 | |
| ГОСТ 34489-2018 | Эскалаторы и пассажирские конвейеры. Правила и методы исследований (испытаний) и измерений. Правила отбора образцов | 01.06.2020 | |
| ГОСТ 33653-2019 | Лифты. Специальные требования безопасности. Вандалозащищенность | 01.06.2020 | |
| ГОСТ 33652-2019 | Лифты. Специальные требования безопасности и доступности для инвалидов и других маломобильных групп населения | 01.06.2020 | |
| ГОСТ 34580-2019 | Лифты. Специальные требования доступности для инвалидов и других маломобильных групп населения при капитальном ремонте и приспособлении зданий и сооружений | 01.06.2020 | |
| ГОСТ 34581-2019 | Лифты. Специальные требования безопасности при установке новых лифтов в существующие здания | 01.06.2020 | |
| ГОСТ 34582-2019 | Лифты. Правила и методы испытаний, измерений и проверок перед вводом в эксплуатацию | 01.06.2020 | |
| ГОСТ 34583-2019 | Лифты. Правила и методы испытаний, измерений и проверок в период эксплуатации | 01.06.2020 |
Химия
| ==================== | |||
| ГОСТ Р 58473-2019 | Классификация опасности химической продукции. Общие требования | 01.06.2022 | |
| ГОСТ Р 58474-2019 | Предупредительная маркировка химической продукции. Общие требования | 01.06.2022 | |
| ГОСТ Р 58475-2019 | Паспорт безопасности химической продукции. Общие требования | 01.10.2022 | |
| ГОСТ 34558-2019 | Методы испытания по воздействию химической продукции на организм человека. Исследование отдаленного нейротоксического действия фосфорорганических соединений при повторном 28-дневном введении | 01.06.2020 | |
| ГОСТ 34556-2019 | Методы испытания по воздействию химической продукции на организм человека. Испытания по оценке кожной сенсибилизации методом изучения реакции региональных лимфатических узлов | 01.06.2020 | |
| ГОСТ 34555-2019 | Методы испытания по воздействию химической продукции на организм человека. Определение токсичности при повторном/многократном воздействии с одновременным определением оценки репродуктивной/эмбриональной токсичности скрининговым методом | 01.06.2020 | |
| ГОСТ 34554-2019 | Методы испытания по воздействию химической продукции на организм человека. Испытания по оценке репродуктивной токсичности двух поколений | 01.06.2020 | |
| ГОСТ 34553-2019 | Методы испытания по воздействию химической продукции на организм человека. Токсикокинетические испытания | 01.06.2020 | |
| ГОСТ Р 58479-2019 | Руководство по применению критериев классификации опасности химической продукции по воздействию на организм. Избирательная токсичность на органы-мишени и/или системы при однократном воздействии | 01.06.2020 | |
| ГОСТ Р 58478-2019 | Руководство по применению критериев классификации опасности химической продукции по воздействию на организм. Опасность при аспирации | 01.06.2020 | |
| ГОСТ Р 58477-2019 | Руководство по применению критериев классификации опасности химической продукции по воздействию на организм. Избирательная токсичность на органы-мишени и/или системы при многократном/продолжительном воздействии | 01.06.2020 | |
| ГОСТ Р 58476-2019 | Руководство по применению критериев классификации опасности химической продукции по воздействию на организм. Сенсибилизирующее действие | 01.06.2020 | |
| ГОСТ Р 58927-2020 | Колонки топливораздаточные. Общие технические условия | 01.10.2020 |
Горнорудное производство
| ==================== | ||
| ГОСТ Р 58585-2019 | Национальный стандарт Российской Федерации. Оборудование горно-шахтное. Общие требования безопасности к системам электроснабжения и управления | 01.04.2020 |
| ГОСТ Р 58493-2019 | Национальный стандарт Российской Федерации. Оборудование горно-шахтное. Машины буропогрузочные. Общие технические требования и методы испытаний | 01.01.2020 |
| ГОСТ Р 58584-2019 | Национальный стандарт Российской Федерации. Горное дело. Пункты переключения в самоспасатели. Требования безопасности. Методы испытаний | 01.04.2020 |
| ГОСТ Р 58718-2019 | Национальный стандарт Российской Федерации. Горное дело. Кабели гибкие шахтные. Методы проверки обеспечения опережающего отключения при раздавливании | 01.07.2020 |
| ГОСТ Р 58717-2019 | Национальный стандарт Российской Федерации. Горное дело. Ремонт и испытание шахтных силовых кабелей. Общие технические требования | 01.07.2020 |
| ГОСТ 34595-2019 | Межгосударственный стандарт. Горное дело. Патроны пневмоимпульсные. Общие технические требования. Методы испытаний | 01.06.2020 |
Электроустановки
| ==================== | ||
| ГОСТ Р 58687-2019 | Взрывоопасные среды. Компетентность персонала организаций, осуществляющих монтаж и первичную проверку электроустановок для применения во взрывоопасных средах. Требования | 01.06.2020 и все ниже |
| ГОСТ Р 58688-2019 | Взрывоопасные среды. Правила оценки соответствия компетентности персонала, осуществляющего монтаж, техническое обслуживание, ремонт и восстановление электроустановок для применения во взрывоопасных средах | |
| ГОСТ Р 58689-2019 | Взрывоопасные среды. Компетентность персонала организаций, осуществляющих проверку и техническое обслуживание электроустановок для применения во взрывоопасных средах. Требования | |
| ГОСТ Р 58690-2019 | Взрывоопасные среды. Компетентность персонала организаций, осуществляющих ремонт и восстановление электроустановок для применения во взрывоопасных средах. Требования | |
| ГОСТ Р 58691-2019 | Взрывоопасные среды. Системы менеджмента качества организаций, осуществляющих монтаж и первичную проверку электроустановок для применения во взрывоопасных средах. Требования | |
| ГОСТ Р 58692-2019 | Взрывоопасные среды. Правила оценки соответствия систем менеджмента качества организаций, осуществляющих монтаж и первичную проверку электроустановок для применения во взрывоопасных средах | |
| ГОСТ Р 58693-2019 | Взрывоопасные среды. Правила оценки соответствия систем менеджмента качества организаций, осуществляющих проверку и техническое обслуживание электроустановок для применения во взрывоопасных средах | |
| ГОСТ Р 58695-2019 | Взрывоопасные среды. Правила оценки соответствия систем менеджмента качества организаций, осуществляющих ремонт и восстановление электроустановок для применения во взрывоопасных средах | |
| ГОСТ Р 58698-2019 | Защита от поражения электрическим током. Общие положения для электроустановок и электрооборудования | |
| ГОСТ Р 58761-2019 | Национальный стандарт Российской Федерации. Здания мобильные (инвентарные). Электроустановки. Технические условия | 01.09.2020 |
Проектирование и строительство
| ==================== | ||
| СП 18.13330.2019 | СП 18.13330.2019. Свод правил. Производственные объекты. Планировочная организация земельного участка (СНиП II-89-80* «Генеральные планы промышленных предприятий») | 18.03.2020 |
| СП 471.1325800.2019 | СП 471.1325800.2019. Свод правил. Информационное моделирование в строительстве. Контроль качества производства строительных работ | 25.06.2020 |
| ГОСТ Р 58754-2019 | Национальный стандарт Российской Федерации. Подмости передвижные с перемещаемым рабочим местом. Технические условия | 01.09.2020 |
| ГОСТ Р 58752-2019 | Национальный стандарт Российской Федерации. Средства подмащивания. Общие технические условия | 01.09.2020 |
| ГОСТ 21.508-2020 | Система проектной документации для строительства. Правила выполнения рабочей документации генеральных планов предприятий, сооружений и жилищно-гражданских объектов | 01.01.2021 |
| ГОСТ Р 21.101-2020 | Система проектной документации для строительства. Основные требования к проектной и рабочей документации | 01.01.2021 |
Сварочное производство
=========================== | ||
ГОСТ Р 58721-2019 | Соединения сварные из сталей марок 10гн2мфа, 15х2нмфа деталей оборудования и трубопроводов атомных энергетических установок. Требования к сварке, наплавке и термической обработке | 01.04.2020 |
| ГОСТ Р 58904-2020 | Сварка и родственные процессы. Словарь. Часть 1. Общие термины | 01.10.2020 |
| ГОСТ Р 58906-2020 | Сварка и родственные процессы. Словарь. Часть 4. Дуговая сварка | 01.10.2020 |
| ГОСТ ISO 9692-2-2020 | Сварка и родственные процессы. Типы подготовки соединений. Часть 2. Сварка дуговая сталей под флюсом | 01.12.2020 |
| ГОСТ ISO 9692-3-2020 | Сварка и родственные процессы. Типы подготовки соединений. Часть 3. Сварка дуговая в инертном газе плавящимся и вольфрамовым электродом алюминия и его сплавов | 01.12.2020 |
| ГОСТ Р ИСО 9692-4-2020 | Сварка и родственные процессы. Рекомендации по подготовке соединений. Часть 4. Плакированные стали | 01.12.2020 |
| ГОСТ ISO 15609-3-2020 | Технические требования и аттестация процедур сварки металлических материалов. Технические требования к процедуре сварки. Часть 3. Электронно-лучевая сварка | 01.12.2020 |
| ГОСТ ISO 15609-5-2020 | Технические требования и аттестация процедур сварки металлических материалов. Технические требования к процедуре сварки. Часть 5. Контактная сварка | 01.12.2020 |
| ГОСТ ISO 14341-2020 | Материалы сварочные. Проволоки и наплавленный металл дуговой сварки плавящимся электродом в защитном газе нелегированных и мелкозернистых сталей. Классификация | 01.12.2020 |
| ГОСТ ISO 14171-2020 | Материалы сварочные. Проволоки сплошного сечения, порошковые проволоки и комбинации проволока/флюс для дуговой сварки под флюсом нелегированных и мелкозернистых сталей. Классификация | 01.12.2020 |
| ГОСТ Р ИСО 18275-2020 | Материалы сварочные. Электроды покрытые для ручной дуговой сварки высокопрочных сталей. Классификация | 01.11.2020 |
| ГОСТ ISO/TR 15608-2020 | Сварка. Руководство по системе группирования металлических материалов | 01.12.2020 |
| ГОСТ ISO 6848-2020 | Дуговая сварка и резка. Электроды неплавящиеся вольфрамовые. Классификация | 01.12.2020 |
| ГОСТ Р ИСО 3580-2020 | Материалы сварочные. Электроды покрытые для ручной дуговой сварки жаропрочных сталей. Классификация | 01.12.2020 |
Нормативы | в Крыму
План 2018 года по разработке строительных стандартов и Сводов правил в рамках программы национальной стандартизации выполнен полностью, акцент в работе был сделан на разработку нормативной документации, касающейся инновационных материалов и технологий. В планах будущего года – актуализация нормативной базы для исключения устаревших материалов и технологий. Об этом сообщил глава Минстроя России Владимир Якушев ,подводя итоги работы в области технического регулирования.В 2018 году разработано 58 проектов сводов правил, 77 проектов национальных и межгосударственных строительных…
Минимальные отступы зданий, строений, сооружений от границ земельных участков СП 42.13330.2016 Градостроительство. Планировка и застройка городских и сельских поселений. Между длинными сторонами жилых зданий следует принимать расстояния (бытовые разрывы): для жилых зданий высотой два-три этажа — не менее 15 м; четыре этажа — не менее 20 м; между длинными сторонами и торцами этих же зданий с окнами из жилых комнат — не менее 10 м. В условиях реконструкции и других сложных градостроительных условиях указанные расстояния могут быть сокращены при соблюдении норм инсоляции, освещенности и…
Строительные нормы и правила (СНиП) — совокупность принятых органами исполнительной власти нормативных актов технического, экономического и правового характера, регламентирующих осуществление градостроительной деятельности, а также инженерных изысканий, архитектурно-строительного проектирования и строительства. Система нормативных документов в строительстве в СССР действовала наряду с системой стандартизации в строительстве, являющейся частью Государственной системы стандартизации, а также с системой стандартизации в рамках СЭВ. С 1995 года СНиПы являлись частным случаем технических…
Государственный стандарт — основная категория стандартов в СССР, сегодня межгосударственный стандарт в СНГ. Принимается Межгосударственным советом по стандартизации, метрологии и сертификации (МГС). В настоящее время являются нормативными неправовыми актами.Стандарты, принятые до 1996 года, являлись нормативно-правовыми актами и поэтому были обязательными для применения.. Для документов, принятых после 1996 года, нормативность сама по себе перестала означать обязательность документа. В настоящее время документ становится обязательным нормативно-правовым актом после регистрации в…
Для повышения качества проектирования, сокращения сроков выполнения работ необходима четкая система нормирования. В настоящее время при значительном количестве актуализированных СНиП и разработанных новых Сводах правил.Их использование при проектировании ограничено отсутствием утвержденного нового Перечня национальных стандартов и сводов правил (частей таких стандартов и сводов правил), в результате применения которых на обязательной основе обеспечивается соблюдение требований Федерального закона «Технический регламент о безопасности зданий и сооружений». Данный Перечень был утвержден…
В последнее время, особенно в связи с организацией СРО все более актуальным становится вопрос сертификации строительных компаний на соответствие требованиями международного стандарта ИСО 9001. В ФЗ «О Саморегулируемых организациях» заложена возможность самим СРО разрабатывать стандарты/регламенты, в том числе включая требования к системе менеджмента качества. Большая доля вероятности, что за основу будет браться именно стандарт ИСО 9001 в отраслевом применении. Перед руководством строительных предприятий встает и будет вставать вопрос — что такое система менеджмента…
В 2009 году строительное лицензирование было отменено. В мировой практике строительства уже давно найдена замена лицензии — сертификат соответствия стандартам ISO 9000 (ГОСТ Р ИСО 9001-2001). Национальный Экспертный Совет по Качеству назвал стандарт ISO 9000 самым популярным в России. Аналитики НЭСК утверждают, что за последние 3 года количество российских строительных компаний, имеющих сертификат ISO 9000, выросло в 5 раз и составляет третью часть от их общего числа. При этом отмечается, что рост популярности сертификата отразится на его стоимости. Прогнозируется, что в…
Настоящий стандарт представляет собой аутентичный текст стандарта ИСО 9001-2000 «Системы менеджмента качества. Рекомендации по улучшению деятельности» и направлен на применение «процессного подхода» при разработке, внедрении и улучшении результативности системы менеджмента качества с целью повышения удовлетворенности потребителей путем выполнения их требований. Три первых раздела стандарта являются «служебными», они не содержат в себе требований. А вот пять остальных разделов – с 4-го по 8-ой – содержат самое главное – требования. Каждый раздел стандарта посвящён…
Все нормы регулируются СНиПом 30-02-97 «Планировка и застройка территорий садоводческих объединений граждан, здания, и сооружения. Нормы проектирования» и СП 11-106-97 «Порядок разработки, согласования, утверждения и состав проектно-планировочной документации на застройку территорий садоводческих объединений граждан».Что можно строить на дачном участке? На каком расстоянии должны быть постройки? На эти и другие вопросы можно найти ответы в приведенных ниже таблицах и графиках.Расстояние между домамиПротивопожарные расстояния между строениями одного участка не нормируются. Отдаленность друг…
I-SNP с LTC-Led продолжают выпускаться, но 2022 год станет годом для просмотра
Когда конференции были основным продуктом развития бизнеса, а не потенциальными источниками заражения COVID-19, квалифицированные медицинские работники часто заполняли комнаты, проводя занятия по институциональным планам для особых нужд (I-SNP).
Согласно этой модели, SNF могут перейти в мир Medicare Advantage, предлагая свои собственные планы страхования бенефициарам, которые проживают в учреждении или нуждаются в уходе на дому.
Для многих SNF I-SNP рассматривались как способ избежать финансового давления, связанного с работой с рядом плательщиков, включая планы управляемого медицинского обслуживания и подотчетные организации медицинского обслуживания. А во время пандемии COVID-19 участие в планах с их интеграцией медицинского обслуживания на местах оказалось как финансово, так и клинически полезным.
Короче говоря, сотрудникиSNF с энтузиазмом восприняли эти планы, хотя существуют и другие варианты оказания помощи, основанной на стоимости, и предостерегает множество предупреждений о том, что модель может быть подходящей не для каждого поставщика.В некотором смысле неудивительно, что количество I-SNP, возглавляемое поставщиками услуг долгосрочного ухода (LTC), выросло на 28% в плановом году на 2021 год, согласно анализу консалтинговой фирмы ATI Advisory.
В частности, количество поддерживаемых LTC I-SNP увеличилось с 76 планов в плановом году 2020 до 98 в 2021 году и выросло с 9% от общего числа предложений I-SNP в 2015 году до 37% в 2021 году.
«Это означает, что на рынке I-SNP рост I-SNP под руководством провайдера LTC опережает рост I-SNP под руководством страховщика», — отмечается в анализе.
Источник: ATI AdvisoryНовые организации, вышедшие на рынок, не стали неожиданностью для Энн Тумлинсон, генерального директора ATI Advisory, но она ожидала, что организации, которые имеют более длительный опыт работы в области I-SNP, упадут. .
«Я думала, что некоторые из более старых, более устоявшихся организаций, у которых есть несколько планов… столкнутся с большим спадом», — сказала она в интервью SNN 17 ноября. «Я думал, что мы просто увидим, что некоторые из этих планов будут свернуты и не будут перевыпущены в следующем году, но в основном это не так.”
Она ожидала некоторого спада по многим причинам, по которым I-SNP, как правило, имеют предупреждающие надписи для SNF: модель сложна, особенно в плане ухода за слабым и уязвимым населением, с меньшим числом учащихся и более простыми способами потерять регистрацию. .
Но, с другой стороны, во время пандемии у них тоже есть свои плюсы.
«Большинство этих планов работают с клиническими моделями, которые намного надежнее, чем то, что вы обычно видели бы в доме престарелых или учреждении для престарелых», — отметил Тамлинсон.«У них есть возможность заплатить за телемедицину и возможность найти практикующую медсестру при первых признаках бедствия и быть тут же. … Если что-то и научило нас тому, что нам нужны действительно сильные клинические модели на местах в этих учреждениях, так это такая ситуация ».
Она была особенно удивлена замедлением темпов роста I-SNP, не управляемых провайдером, особенно по сравнению с темпами роста тарифных планов, управляемых провайдером. Первый вырос на 10 с 2020 по 2021 год; последняя увеличилась на 14 планов одновременно.
«Все поставщики готовы. Они преданы делу, это своего рода мой вывод из всего этого», — сказал Тамлинсон SNN.
По крайней мере, это вывод для текущего набора данных. Но Тумлинсон признал, что во многих случаях планы, запускаемые в 2021 году, позволили бы начать процесс задолго до пандемии, что сделало 2022 год годом, за которым нужно наблюдать. Запуск I-SNP может занять от нескольких месяцев до более года, и даже в этом случае достижение стабильности может занять некоторое время, как отметил один из основных административных партнеров I-SNP в интервью 2019 года.
«Нам нравится, чтобы первый год наших планов был стабильным. Это означает, что план нарушается даже на весь год, за что, кстати, большинство планов MA были бы убиты », — сказал тогда SNN президент и генеральный директор AllyAlign Уилл Сондерс. «Затем мы хотим, чтобы второй год был тем, что мы называем жизнеспособностью, то есть вы начинаете приносить прибыль. Тогда 3-й год — это то, что мы называем оптимизацией, когда вы действительно узнали рычаги успеха, ваше учреждение это понимает и вы действительно добиваетесь превосходных результатов для своих участников и их семей.”
Пандемия, вероятно, покончит с этими временными рамками или сильно нарушит их, о чем ранее в этом году сообщили SNN другие планы по запуску I-SNP. Но это не обязательно означает, что модель в мире ОЯТ обречена; вместо этого на каком-то уровне он может быть поставлен на паузу.
Тем не менее, Тумлинсон считает, что 2022 год будет еще более важным годом для изучения с точки зрения предложений I-SNP. Что касается показателей, которые нужно отслеживать для оценки работоспособности I-SNP, сторонние наблюдатели могут отслеживать количество планов, регистрацию, премиальные сборы, личные расходы и дополнительные льготы, но, по словам Тумлинсона, основным показателем является зачисление.
Январь и февраль 2021 года, вероятно, предоставят больше всего информации, когда Центры услуг Medicare и Medicaid (CMS) опубликуют свой файл регистрации для I-SNP, пояснила она. Любой, кто работает в рамках одного из планов, будет следить за частотой госпитализаций и инфекциями COVID-19 среди зачисленного населения.
Но когда дело доходит до оценки влияния COVID-19 на интерес к I-SNP со стороны поставщиков долгосрочного ухода, ключевой момент будет через год, сказал Тамлинсон.
«Я определенно буду задерживать дыхание на 2022 год… и все же я сохраняю оптимизм», — сказала она SNN. «Я замечаю пару тенденций: во-первых, мы можем не увидеть новых организаций в 2022 году, но я думаю, что мы все равно увидим, что существующие организации будут предлагать больше планов. Это количество планов, а не организации, владеющие планами. Некоторые из организаций, которые уже были в этой сфере, фактически увеличили количество планов, которые они предлагали с 2020 по 2021 год, несмотря на все препятствия.”
Применение глубокого обучения для классификации правильных и неправильных генотипов SNP на основе конвейеров секвенирования полногеномной ДНК
Наборы данных
Данные включают считывание полногеномной последовательности ДНК четырех традиционных датских быков красного молочного скота. Образцы секвенировали на платформе Illumina HiSeq2000 с длиной считывания парных концов 100 п.н. с размером вставки 300 п.н. Общее количество необработанных считываний, сгенерированных для одного животного, варьировалось от 249 478 818 до 290 364 464.Это привело к среднему охвату генома в 10 раз. Кроме того, эти быки были генотипированы с использованием набора Illumina BovineHD BeadArray, содержащего 777 962 SNP. Затем эти данные были использованы для составления следующих подмножеств:
Набор обучающих данных, используемый для построения классификатора на основе DL, состоял из трех (из четырех) животных.
Набор данных проверки, используемый в качестве независимых входных данных для проверки качества классификации, включал данные от четвертого животного с SNP NGS, идентифицированными на основе того же конвейера, что и в наборе данных для обучения.
Вызов SNP
Конвейер для вызова SNP включал выравнивание по эталонному геному, выполненное с использованием BWA-MEM (Li and Durbin 2009) со следующими параметрами по умолчанию: длина семени 19, оценка совпадения 1, несоответствие штраф 4, штраф за открытие промежутка 6 и штраф за расширение промежутка 1. Обработка после выравнивания включала преобразование файлов в формате SAM в двоичный (BAM) формат, индексацию данных и маркировку дубликатов PCR.Этот шаг был выполнен с использованием Picard (https://broadinstitute.github.io/picard/) и SAMtools. Предварительный вызов был реализован с помощью пакета GATK и включал локальное повторное выравнивание вокруг INDEL с помощью инструментов GATK RealignerTargetCreator и IndelRealigner с последующей повторной калибровкой показателя качества с помощью инструментов BaseRecalibrator и PrintReads. Для фактического вызова варианта использовался инструмент UnifiedGenotyper из пакета GATK. Обратите внимание, что указанный выше конвейер был запущен только для тех 772 173 геномных позиций SNP, которые также были генотипированы с помощью Illumina BovineHD BeadArray и были определены в SNPchiMp (Nicolazzi et al.2015) база данных.
Правильное и неправильное определение SNP
После исключения отсутствующих генотипов общее количество проанализированных полиморфных локусов для каждого из быков составило 764 446, и их сравнивали между результатами NGS и микрочипами. Правильные SNP относятся к полному совпадению генотипов, оцененных обеими технологиями; неверные SNP были определены как несовпадения, включающие хотя бы один аллель. В случае мультиаллельных SNP, когда в выходных данных NGS вызывались два или более альтернативных аллеля, мы проверяли, допускает ли хотя бы один из этих аллелей совпадение с генотипом, вызванным микрочипом.
Объясняющие переменные
Следующие объясняющие переменные из стандартного вывода VCF были рассмотрены для классификации:
Вероятность неправильно названного альтернативного аллеля (QUAL): QUAL = — 10log 10 ( P 1 ), где P 1 обозначает вероятность того, что идентифицированный альтернативный аллель является неправильным обнаружением.
Условная вероятность неправильно названного альтернативного аллеля (GQ): GQ = — 10log 10 ( P 2 ), где P 2 обозначает вероятность того, что идентифицированный генотип является неправильным обнаружением, обусловленным позиция полиморфна.
Глубина секвенирования в полиморфном сайте в совокупности для всех четырех секвенированных индивидов (DP).
Глубина секвенирования в полиморфном сайте для данного человека (DP2).
Закодированный генотип (CALL): \ (\ mathrm {CALL} = \ left \ {\ \ begin {array} {c} 1 \ kern0.5em \ mathrm {for} \ kern0.5em ./. \\ {} \ kern0.75em 2 \ kern0.5em \ mathrm {для} \ kern0.5em 0/0 \\ {} \ kern0.75em 3 \ kern0.5em \ mathrm {for} \ kern0. 5em 0/1 \\ {} \ kern1em 4 \ kern0.5em \ mathrm {for} \ kern0.5em 1/1 \ \ end {array} \ right. \).
Категориальная переменная была построена на основе трех базовых ссылок ниже SNP.
Категориальная переменная была построена на основе трех базовых ссылок перед SNP.
Последние три переменные были включены в моделирование, чтобы уловить потенциальные ошибки секвенатора, специфичные для флуоресценции определенных генотипов (CALL) или последовательности, близко соседней с полиморфным сайтом.
Алгоритмы глубокого обучения
Алгоритмы глубокого обучения были реализованы через интерфейс Keras (Chollet 2015) с библиотекой TensorFlow (Abadi et al. 2015) в Python 3.7.7 на персональном компьютере под управлением Windows 10 с Intel Core i5 -3210M CPU 2.50 ГГц (2 ядра и кэш-память 3 МБ), 8 ГБ ОЗУ и 120 ГБ SSD. Перед реализацией алгоритма все количественные объясняющие переменные (QUAL, GQ, DP и DP2) были преобразованы в стандартное нормальное распределение. Категориальные независимые переменные (базовое трио до и после SNP, а также CALL) кодировались в горячем режиме. Сетевая архитектура, лежащая в основе простого алгоритма (NAÏVE), состояла из восьми последовательно соединенных уровней с постепенно уменьшающимся числом параметров и коэффициентом отсева, равным 0.{- {x} _ {ij}}} \) использовался как активация последнего слоя. Алгоритм Adam (Kingma and Ba 2014), реализующий подход стохастического градиентного спуска, использовался для исследования функции правдоподобия, а бинарная кроссентропия функция потерь использовалась для количественной оценки качества классификации, применяемой к набору обучающих данных. Реализация Keras представлена на рис. 1.
Рис. 1Схема реализации алгоритма глубокого обучения NAÏVE для классификации SNP в Keras
Чтобы смягчить дисбаланс в количестве классов, наблюдаемый в наборе обучающих данных, приведенное выше Наивный алгоритм был изменен путем предварительного наложения различных весов для правильного и неправильного класса SNP при вычислении метрики потерь.Следуя рекомендациям онлайн-руководства TensorFlow, вес для правильного класса SNP был оценен как \ (\ frac {1} {2} \ cdotp \ frac {N _ {\ mathrm {trueSNP}} + {N} _ {\ mathrm {falseSNP}}} {N _ {\ mathrm {trueSNP}}} \) и для неправильного класса SNP как \ (\ frac {1} {2} \ cdotp \ frac {N _ {\ mathrm {trueSNP}} + {N } _ {\ mathrm {falseSNP}}} {N _ {\ mathrm {falseSNP}}} \), где N представляет количество SNP, представляющих соответствующий класс. Этот алгоритм называется ВЗВЕШЕННЫМ. Еще одна модификация алгоритма NAÏVE была реализована в рамках построения обучающего набора данных.В частности, случайным образом выбирались SNP для неправильного класса с заменой из пула всех неверных SNP. Это привело к большему количеству SNP, представляющих неправильный класс SNP. В алгоритме OVERSAMPLED30 количество неверных SNP равнялось 30% от количества правильных SNP; в алгоритме OVERSAMPLED60 количество неверных SNP было равно 60% от количества правильных SNP, а в алгоритме OVERSAMPLED100 оба класса были представлены равным количеством SNP.
Метрики качества классификации
Классификационный подход включает следующие категории:
Истинно положительный (TP) определяется как ситуация, когда неверный SNP был классифицирован как неправильный.
Ложноотрицательный (FN) определяется как ситуация, когда неверный SNP был классифицирован как правильный.
Истинно отрицательный (TN) определяется как ситуация, когда правильный SNP был классифицирован как правильный.
Ложное срабатывание (FP) определяется как ситуация, когда правильный SNP был классифицирован как неправильный.
На основе этих категорий для количественной оценки качества классификаторов использовались две сводные статистики. Метрика F1 задается как \ (F1 = \ frac {2 \ mathrm {TP}} {2 \ mathrm {TP} + \ mathrm {FN} + \ mathrm {FP}} \), а метрика SUMSS задается как \ (S \ mathrm {UMSS} = \ frac {\ mathrm {TN}} {\ mathrm {TN} + \ mathrm {FP}} + \ frac {\ mathrm {TP}} {\ mathrm {TP} + \ mathrm { FN}} \).
Оценка вероятности отсечения
Для каждого проанализированного SNP исходный результат последнего уровня представляет собой вероятность того, что SNP является правильным, что является результатом функции активации сигмоида. При присвоении класса истинно / ложно по умолчанию применяется порог вероятности 0,5. Для каждого из реализованных алгоритмов (NAÏVE, WEIGHTED и три OVERSAMPLED), в дополнение к этому стандартному порогу, значения отсечения вероятности были также оценены на основе оптимизации показателей F1 или SUMSS соответственно с использованием пакета cutpointR (Thiele and Hirschfeld 2020). реализована в R.Вкратце, в этом пакете отдельно для каждого из пяти алгоритмов выполняется многократная выборка подмножества данных с заменой из исходного обучающего набора SNP. Для каждой такой подвыборки граница вероятности представлена значением, которое дает наивысшую метрику F1 / SUMSS. Окончательная оценка — это среднее значение отсечки для всех выборок начальной загрузки. Чтобы проверить надежность оценок отсечения по отношению к исходным данным, оценки отсечения были проверены путем их оценки на основе 20 подвыборок наших исходных обучающих данных.
Новая технология генотипирования SNP Target SNP-seq и ее применение в генетическом анализе сортов огурцов
Vignal, A., Milan, D., SanCristobal, M. & Eggen, A. Обзор SNP и др. типы молекулярных маркеров и их использование в генетике животных. Genet. Selection Evolution, 34,3 34 , 275–305 (2002).
CAS Статья Google ученый
Перкель, Дж. Ошибка: генотипирование SNP: шесть технологий, совершивших революцию. Nat. Методы 5 , 447–453 (2008).
CAS Статья Google ученый
Ляо, П. Я. и Ли, К. Х. От SNP к функциональному полиморфизму: понимание приложений биотехнологии. Biochemical Eng. J. 49 , 149–158 (2010).
CAS Статья Google ученый
Йем, I. Текущие достижения и перспективы вирусной устойчивости садовых культур. Садоводство, Окружающая среда, Биотехнологии. 57 , 113–122, https://doi.org/10.1007/s13580-016-0105-x (2016).
CAS Статья Google ученый
Шавруков Ю.В. Сравнение применения маркеров SNP и CAPS в генетических исследованиях пшеницы и ячменя. Завод БМК. Биол. 16 , 11 (2016).
Артикул Google ученый
Томсон, М. Дж. Высокопроизводительное генотипирование SNP для ускорения улучшения сельскохозяйственных культур. Завод. Порода. Biotechnol. 2 , 195–212 (2014).
Артикул Google ученый
Тиан, Х. Л. и др. . Разработка кукурузы SNP3072, массива SNP, совместимого с высокой пропускной способностью, для идентификации китайских сортов кукурузы по отпечаткам ДНК. Мол. Порода. 35 , 136 (2015).
Артикул Google ученый
Shirasawa, K., Monna, L., Kishitani, S. & Nishio, T. Однонуклеотидный полиморфизм в случайно выбранных генах среди сортов риса японская (Oryza sativa L.), идентифицированных с помощью ПЦР-RF-SSCP. Dna Res. 11 , 275–283 (2004).
CAS Статья Google ученый
Лю Л. и др. . Режим высокопроизводительной идентификации сортов пшеницы по маркерам SNP. Журнал Triticeae Crops (2018).
Kuang, M. et al. . Разработка основного набора маркеров SNP для идентификации сортов хлопчатника высокогорных районов Китая. J. Integr. Сельское хозяйство 15 , 954–962 (2016).
Артикул Google ученый
Wei, Z. Y., Hui Hui, L. I., Jun, L. I. & Yasir. Точная идентификация сортов по нуклеотидным полиморфизмам и установление сканируемых идентификаторов сортов зародышевой плазмы сои. Acta Agronomica Sinica (2018).
Лю В. и др. . Влияние выбора формы плодов на генетическую структуру и разнообразие, выявленное при генотипировании идеальных SNP по всему геному в баклажанах. Мол. Порода. 39 , 140, https://doi.org/10.1007/s11032-019-1051-y (2019).
Артикул Google ученый
Du, H. et al. . Целевое секвенирование выявляет генетическое разнообразие, структуру популяции, маркеры ядра-SNP и локусы, связанные с формой плода, у разновидностей перца. Завод БМК. Биол. 19 , 578, https://doi.org/10.1186/s12870-019-2122-2 (2019).
CAS Статья PubMed PubMed Central Google ученый
Jung, J. K., Park, S. W., Liu, W. Y. & Kang, B. C. Обнаружение однонуклеотидного полиморфизма в маркерах Capsicum и SNP для идентификации сортов. Euphytica 175 , 91–107 (2010).
CAS Статья Google ученый
Ван, К. К. и др. . Разработка экономичного нового метода для быстрого одновременного генотипирования пяти общих однонуклеотидных полиморфизмов гена нейротрофического фактора головного мозга (BDNF) с помощью системы устойчивых к амплификации тетрапраймеров мутаций. Внутр. J. Methods Psychiatr. Res. 24 , 235 (2015).
Артикул Google ученый
Семагн, К., Бабу, Р., Херн, С. и Олсен, М.Генотипирование по однонуклеотидному полиморфизму с использованием Kompetitive Allele Specific PCR (KASP): обзор технологии и ее применения для улучшения сельскохозяйственных культур. Мол. Порода. 33 , 1–14 (2014).
CAS Статья Google ученый
Лоу, Ю. Л., Ведрен, С. и Лю, Дж. Геномная технология с высокой пропускной способностью в исследованиях и клиническом лечении рака груди. Развитие генетических эпидемиологических исследований. Breast Cancer Res. BCR 8 , 209–209 (2006).
Артикул Google ученый
Law, W. D., Fogarty, E. A., Vester, A. & Antonellis, A. Полногеномная оценка консервативных аллелей SNP выявляет панель регуляторных SNP, относящихся к периферическому нерву. BMC Genomics 19 , 311–311, https://doi.org/10.1186/s12864-018-4692-z (2018).
CAS Статья PubMed PubMed Central Google ученый
Мамедов Дж., Аггарвал Р., Буйярапу Р. и Кумпатла С. SNP-маркеры и их влияние на селекцию растений. Внутр. Дж. Плант. Геномика, 2012, 2012 , 728398 (2012).
Google ученый
Мардис, Э. Р. Влияние технологии секвенирования следующего поколения на генетику. Тенденции генетики: TIG 24 , 133–141, https://doi.org/10.1016/j.tig.2007.12.007 (2008).
CAS Статья Google ученый
Менг, К. и др. . Разработка маркеров SNP с использованием технологии RNA-seq и тетрапраймера ARMS-PCR в сладком картофеле. J. Integr. Сельское хозяйство 16 , 464–470 (2017).
Артикул Google ученый
Чен, К. и др. . Новая трехэтапная мультиплексная ПЦР для генотипирования SNP с секвенированием следующего поколения. Анал. Биоанал. Chem. 408 , 4371–4377, https: // doi.org / 10.1007 / s00216-016-9536-6 (2016).
CAS Статья Google ученый
Онда, Ю., Такахаги, К., Симидзу, М., Иноуэ, К. и Мочида, К. Мультиплексное целевое секвенирование ампликонов ПЦР (MTA-Seq): простое, гибкое и универсальное генотипирование SNP с помощью Секвенирование ампликонов с высокой степенью мультиплексирования с помощью ПЦР. Frontiers in Plant Science 9 , https://doi.org/10.3389/fpls.2018.00201 (2018).
Ян С. и др. . Платформа маркерного генотипирования следующего поколения (AmpSeq) в гетерозиготных культурах: тематическое исследование селекции с помощью маркеров в виноградной лозе. Horticulture Research 3 , 16002.
Guo, Z. et al . Разработка панелей множественных маркеров SNP, доступных для селекционеров, путем генотипирования с помощью целевого секвенирования (GBTS) кукурузы. Мол. Порода. 39 , 37, https://doi.org/10.1007/s11032-019-0940-4 (2019).
Артикул Google ученый
Редди, П. П. Огурец. (2016).
Lv, J. et al. . Генетическое разнообразие и популяционная структура огурца (Cucumis sativus L.). PLoS One 7 , e46919 (2012).
CAS ОБЪЯВЛЕНИЯ Статья Google ученый
Мяо, Х., Чжан, С. П., Син-Фанг, Г. У., Ван, Й. и Чен, Л. Анализ генетического разнообразия и конструирование по отпечаткам пальцев основных сортов китайского огурца с помощью полиморфизмов с повторением простой последовательности. Журнал генетических ресурсов растений (2014).
Chandel, G. & Banerjee, S. Понимание роли генов-кандидатов, связанных с гомеостазом металлов, в поглощении, транспорте и перераспределении Fe / Zn в рисе с использованием полуколичественной ОТ-ПЦР. J. Plant. Мол. Биол. Biotechnol. 2 , 33–46 (2011).
Google ученый
Cavagnaro, P. F. et al. . Полногеномная характеристика простых повторов последовательности у огурца (Cucumis sativus L.). BMC Genomics 11 , 569 (2010).
Артикул Google ученый
Huang, S. et al. . Геном огурца Cucumis sativus L. Nat. Genet. 41 , 1275 (2009).
CAS Статья Google ученый
Ци, Дж. и др. . Карта геномных вариаций дает представление о генетической основе одомашнивания и разнообразия огурцов. Nature Genetics 45 , 1510, https://doi.org/10.1038/ng.2801, https://www.nature.com/articles/ng.2801#supplementary-information (2013).
Ян, Дж. и др. . Target SSR-Seq: новая технология генотипирования SSR, связанная с идеальными SSR в генетическом анализе сортов огурца. Фронт. Растение. Sci. 10 , 531, https://doi.org/10.3389/fpls.2019.00531 (2019).
Артикул PubMed PubMed Central Google ученый
Li, H. & Durbin, R. Быстрое и точное согласование коротких считываний с помощью преобразования Барроуза – Уиллера. Биоинформатика 25 , 1754–1760, https://doi.org/10.1093/bioinformatics/btp324 (2009).
CAS Статья PubMed PubMed Central Google ученый
Де Сумма, С. и др. . Жесткая фильтрация GATK: настраиваемые параметры для улучшения варианта, требующего секвенирования данных целевой генной панели следующего поколения. BMC Bioinforma. 18 , 119, https://doi.org/10.1186/s12859-017-1537-8 (2017).
CAS Статья Google ученый
Паллави, Х. М., Гоуда, Р., Вишванат, К., Шадакшари, Ю. Г. и Бханупракаш, К. Идентификация маркеров SSR для тестирования гибридности и генетической чистоты семян подсолнечника (Helianthus annuus L.). Seed Sci. Technol. 34 , 59–66 (2011).
Google ученый
Ботштейн Д., Уайт Р. Л., Сколник М. и Дэвис Р. В. Построение карты генетического сцепления у человека с использованием полиморфизмов длины рестрикционных фрагментов. Am. J. Hum. Genet. 32 , 314–331 (1980).
CAS PubMed PubMed Central Google ученый
Хантер П. Числовой индекс дискриминационной способности систем набора текста: применение индекса разнообразия Симпсона. Журнал клинической микробиологии 26 (1988).
Притчард, Дж. К., Стивенс, М. и Доннелли, П. Вывод структуры популяции с использованием данных мультилокусного генотипа. Генетика 155 (2000).
Эванно, Г., Регнаут, С. и Годе, Дж. Определение количества групп людей с помощью программного обеспечения СТРУКТУРА: исследование моделирования. Мол. Ecol. 14 , 2611–2620 (2005).
CAS Статья Google ученый
FUJII, H. et al. . Минимальный маркер: алгоритм и компьютерная программа для идентификации минимальных наборов дискриминирующих маркеров ДНК для эффективной идентификации сорта. J. Bioinforma. Computational Biol. 11 , 1250022, https://doi.org/10.1142/s0219720012500229 (2013).
Артикул Google ученый
УПОВ. Возможное использование молекулярных маркеров при исследовании отличимости, однородности и стабильности (DUS). УПОВ Женева, Швейцария (2011).
Рольф, Ф. Дж. NTSYSpc .: Система числовой таксономии и многомерного анализа. Версия 2.1. Exeter Software, Сетокет, Нью-Йорк . (2000).
Патер, Дж. А. и др. . Распространенный вариант CLDN14 вызывает резкую предлингвальную сенсоневральную тугоухость в нескольких семьях из-за эффекта основателя. Hum. Genet. 136 , 107–118 (2017).
CAS Статья Google ученый
Wongsaprom, C. и др. . Два интрогрессированных локуса количественных признаков придают широкий спектр устойчивости к бластной болезни в генетическом фоне сорта RD6 тайский клейкий жасминовый рис. Урожай полевой. Res. 119 , 245–251 (2010).
Артикул Google ученый
Маманова Л. и др. . Стратегии целевого обогащения для секвенирования следующего поколения. Nature Methods 7 , 111, https: // doi.org / 10.1038 / nmeth.1419, https://www.nature.com/articles/nmeth.1419#supplementary-information (2010).
Tsuchihashi, Z. & Dracopoli, N.C. Прогресс в высокопроизводительных методах генотипирования SNP. Pharmacogenomics J. 2 , 103, https://doi.org/10.1038/sj.tpj.6500094 (2002).
CAS Статья Google ученый
Лю, Дж. и др. . Усовершенствованный метод конструирования аллель-специфических праймеров ПЦР для анализа маркеров SNP и его применение. Завод. Методы 8 , 34, https://doi.org/10.1186/1746-4811-8-34 (2012).
CAS Статья PubMed PubMed Central Google ученый
Liu, Q. et al. . Шаги по обеспечению точности вызова генотипа и SNP из данных секвенирования Illumina. BMC Genomics 13 , S8, https://doi.org/10.1186/1471-2164-13-S8-S8 (2012).
Артикул PubMed PubMed Central Google ученый
Реннер, С.С., Шефер, Х. и Коциан, А. Филогенетика Cucumis (Cucurbitaceae): Огурец (C. sativus) принадлежит к азиатско-австралийской кладе, удаленной от дыни (C. melo). Bmc Evolut. Биол. 7 , 58 (2007).
Артикул Google ученый
Emanuelli, F. et al. . Генетическое разнообразие и популяционная структура, оцененная с помощью маркеров SSR и SNP в большой коллекции зародышевой плазмы винограда. Завод БМК.Биол. 13 , 39–39 (2013).
CAS Статья Google ученый
Cericola, F. et al. . Структура популяции и разнообразие баклажанов из Азии и Средиземноморского бассейна. PLoS one 8 , e73702 – e73702, https://doi.org/10.1371/journal.pone.0073702 (2013).
CAS ОБЪЯВЛЕНИЯ Статья PubMed PubMed Central Google ученый
Чен, Х. М. и др. . Сходство и различие между Китайскими методиками тестирования на ООС и руководящими принципами УПОВ по фаленопсису. Рефераты по китайскому садоводству (2013).
Tommasini, L. et al. . Разработка мультиплексных маркеров простых повторов последовательности (SSR) в дополнение к испытаниям на отличимость, однородность и стабильность сортов рапса (Brassica napus L.). Теор. Прил. Genet. 106 , 1091–1101 (2003).
CAS Статья Google ученый
Кришнан, С. и др. . Интеграция фонового анализа на основе маркеров с отбором на переднем плане для идентификации рекомбинантов с превосходной устойчивостью к бактериальному ожогу в рисе басмати . Vol. 127 (2008).
Jeong, S.C. & Saghai Maroof, M.A. Обнаружение и генотипирование SNP, тесно связанных с двумя локусами устойчивости к болезням, Rsv1 и Rsv3, сои. Завод. Порода. 123 , 305–310, https://doi.org/10.1111/j.1439-0523.2004.00981.x (2004).
CAS Статья Google ученый
Фанг, Д. Д., Сяо, Дж., Канчи, П. К. и Кантрелл, Р. Г. Новый гаплотип SNP, связанный с геном устойчивости к синей болезни у хлопка (Gossypium hirsutum L.). Теор. Прил. Genet. 120 , 943–953, https://doi.org/10.1007/s00122-009-1223-y (2010).
CAS Статья Google ученый
Jänsch, M. et al. . Идентификация SNP, связанных с восемью локусами устойчивости к болезням яблони. Мол. Порода. 35 , 45, https://doi.org/10.1007/s11032-015-0242-4 (2015).
CAS Статья Google ученый
Wen, C. et al. . Тонкое генетическое картирование гена устойчивости к пятнистости листьев cca-3 у огурца Cucumis sativus L. Theor. Прил. Genet. 128 , 2495–2506, https: // doi.org / 10.1007 / s00122-015-2604-z (2015).
CAS Статья PubMed PubMed Central Google ученый
Биологические ограничения на SNP GWAS на предполагаемых порогах значимости выявляют дополнительные локусы ИМТ
Существенные изменения:
1) Это не первое исследование, демонстрирующее, что биологические аннотации в сочетании с более ослаблением пороговых значений статистических ассоциаций из GWAS «спасают» истинные ассоциации в области ожирения.В качестве иллюстрации, Meyre et al., 2009, спасли скромный сигнал ассоциации GWAS стадии 1 для крайнего ожирения в гене NPC1, используя стратегию гена-кандидата, и ассоциация была воспроизведена на стадии 2. Удар ожирения GWAS в NPC1 был недавно подтвержден. как значимый для всего генома сигнал для ИМТ в большом метаанализе (Turcot et al., 2018). Wang et al. (Diabetes 2019) продемонстрировали сильное обогащение положительных ассоциаций с ИМТ для SNP, расположенных в генах синдромального ожирения или рядом с ними. Авторы могут обсудить эти и другие отчеты в литературе.
Рецензент поднимает хороший вопрос и говорит о возможности применения единого геномного подхода, который мы использовали. В ответ на эти моменты мы теперь ссылаемся на предыдущие работы в этом контексте использования биологических аннотаций для спасения подпороговых сигналов. Мы воспользовались предложением включить ключевой пример исследования Meyre et al., 2009 в качестве введения в идею подпорогового майнинга SNP вместе со ссылкой на Turcot et al. бумага, которая воспроизводит эту ассоциацию.Мы также добавили дополнительное исследование Wang et al., 2016, поскольку оно в значительной степени относится к этой статье в плане использования эпигеномных данных для получения биологически значимых подпороговых сигналов от GWAS.
2) Раздел о сильных сторонах и ограничениях добавил бы много в Обсуждение.
Мы согласны с тем, что раздел о сильных сторонах и ограничениях является отличным дополнением к рукописи. Хотя ранее мы включили ряд моментов, касающихся ограничений этого исследования, мы признаем, что требуется более полная разбивка.Мы добавили гораздо более подробное обсуждение уровней классификации, отметив относительно низкую чувствительность, а также подчеркнув высокую точность и специфичность. Мы считаем, что этот компромисс является приемлемым результатом, учитывая, что положительные прогнозы в значительной степени достигли общегеномного значения. В соответствии с комментарием №4 мы также отметили в наших ограничениях отсутствие более разнообразного набора типов клеток, хотя адипоциты и нейроны гипоталамуса являются главными кандидатами, ясно, что есть другие соответствующие типы клеток, которые могут быть использованы в таком исследовании. ; действительно, они могут стать предметом будущего исследования в этом отношении.
3) Биологические критерии (доступность хроматина / прямой контакт с промоторами генов), использованные в этом исследовании, очень оригинальны, но, чтобы быть более исчерпывающими, для выбора большего количества SNP могли использоваться дополнительные биологические критерии (перечисленные в Li and Meyre, Int J Obes 2013 в разделе «Анализ GWAS, основанный на гипотезах»). Это можно признать ограничением исследования.
Мы согласны с тем, что всегда можно использовать больше критериев и ограничений. Наше намерение состояло в том, чтобы наблюдать, достаточно ли этой уникальной и относительно простой комбинации состояния хроматина и физических контактов между задействованными прокси-SNP и промоторами генов для спасения подпороговых SNP.Мы не сомневаемся, что дополнительные фильтры могут в некоторой степени улучшить прогнозы, особенно при более мягких пороговых значениях p , поэтому мы выделили этот момент в нашем Обсуждении, представив возможность использования дополнительных критериев в будущем исследовании для улучшения качество прогнозов.
4) Ткани, включенные в исследование (адипоциты и гипоталамус), чрезвычайно важны в контексте генетической предрасположенности к ожирению. Однако исследования генетических ассоциаций также подчеркнули важную роль других тканей в энергетическом балансе (например,грамм. бета-клетки, печень, мышцы, желудок… см. Locke et al., 2015, и Pigeyre et al., Clin Sci 2016). Отсутствие исследования всех тканей, потенциально значимых для ожирения, можно признать еще одним ограничением исследования.
Мы согласны с тем, что всегда можно использовать больше критериев и ограничений. Наше намерение состояло в том, чтобы наблюдать, достаточно ли этой уникальной и относительно простой комбинации состояния хроматина и физических контактов между задействованными прокси-SNP и промоторами генов для спасения подпороговых SNP.Мы не сомневаемся, что дополнительные фильтры могут в некоторой степени улучшить прогнозы, особенно при более мягких пороговых значениях p , поэтому мы выделили этот момент в нашем Обсуждении, представив возможность использования дополнительных критериев в будущем исследовании для улучшения качество прогнозов.
5) В то время как Speliotes et al., 2010 и Yengo et al., 2018 GWAS для ИМТ были выполнены в популяциях европейского происхождения, исследование Locke et al., 2015 включало многоэтническую популяцию.Анализировали ли авторы сводную статистику GWAS среди населения Европы в исследовании Locke et al., 2015? Если да, они могут предоставить более подробную информацию о том, что они сделали, в разделе «Материалы и методы». Если нет, я думаю, что использование сводной статистики GWAS может добавить некоторую неоднородность, и я рекомендую сосредоточить анализ на европейском населении в исследовании Locke et al., 2015.
Спасибо, что заметили это упущение. Мы использовали результаты европейского происхождения во всех проанализированных исследованиях.Мы обновили Материалы и методы, чтобы отразить этот момент.
6) Организация данных и анализа. Любопытно, что эпигенетические данные, доказательства и анализ данных Capture-C и ATAC-seq находятся в материалах и методах, а не в результатах. Комбинация методов и данных несколько неполна, возможно, из-за их необычного расположения в рукописи. Они являются ключевыми для общего исследования и должны быть в разделе «Результаты». По мере того, как я читал рукопись, мое любопытство росло по поводу характера доказательств связи фланговых и регуляторных SNP с ранее сообщенными целевыми SNP — каков источник данных, какова природа доказательств — только чтобы узнать, что они есть в Материалах и методах, хотя и не полностью.Каковы доказательства наличия открытого хроматина в их мишенях и в контрольных генах, которые наводили на мысль в ранних исследованиях GWAS?
Наше намерение с этой формой презентации состояло в том, чтобы облегчить повествовательный поток и посвятить большую часть раздела результатов процессу, включающему используемый ретроспективный анализ, а обработку эпигенетических данных поместить в раздел материалов и методов. Однако, учитывая комментарий рецензента, мы согласны с тем, что дополнительные сведения должны быть указаны в разделе «Результаты».Поэтому мы добавили дополнительную информацию об этих наборах данных в раздел «Результаты», оставив подробные сведения о том, как эти данные были созданы и обработаны в Материалах и методах. Данные, связанные с нейронами гипоталамуса, были ранее получены и проанализированы в отдельном нашем исследовании, которое мы сейчас цитируем. Кроме того, мы включаем конкретные номера пиков ATAC-seq, а также количество взаимодействий для наших трех библиотек адипоцитов.
7) Дизайн, анализ и презентация исследования.Блок-схема поможет читателям понять последовательность задач и поможет авторам с организацией результатов.
Учитывая сложность этого анализа, мы полностью согласны с этим предложением. Чтобы облегчить понимание, мы добавили блок-схему, описывающую используемые конвейеры (см. Рисунок 3). Блок-схема, которую мы включаем, использует данные BMI за 2010-2015 гг., Чтобы проиллюстрировать наш подход, и включает числа, помогающие описать каждый этап вычислений и количество SNP и локусов, встречающихся на каждом этапе этого анализа.
8) Итог. Ключевой вопрос — в чем преимущество предлагаемого метода? Чего не хватает — простого утверждения о том, сколько суггестивных локусов стали значимыми позже, а сколько — нет, в ходе обычной работы, и параллельно, сколько эпигенетических совпадений с суггестивных локусов стали значимыми позже, а сколько — нет? То есть в чем количественное преимущество нового анализа. Примечательно, что они не показывают цифры для этого, казалось бы, простого и центрального вопроса.Удивительно, но этого пункта нет даже в аннотации; 4-е и 5-е предложения, в которых рассматриваются ключевые результаты, не содержат четких указаний на эти моменты.
Мы принципиально решили не выделять эти числа за пределами различных цифр из-за множества условий, на которых мы работали. Это было сделано для того, чтобы избежать восприятия, что мы каким-то образом выбираем лучший признак, год и интервал значений P , учитывая дисперсию этих чисел, что потенциально сбивает с толку или вводит читателя в заблуждение с самого начала.Тем не менее, мы признаем, что такие числа в тексте были бы полезны, чтобы читателю не приходилось постоянно обращаться к рисункам и таблицам. Поэтому мы добавили такие числа в конце каждого соответствующего раздела, используя подсчеты в пределах согласованного интервала 5 x 10 -8 ≤ P <5 x 10 -5 .
9) Хроматиновые доказательства. Доказательства основаны на двух клеточных линиях — адипоцитах, происходящих из МСК, и гипоталамо-подобных нейронах, происходящих из ESC.Оба являются соответствующими типами клеток in vivo. Но помимо этой функциональной связи, не приводится никакого другого обоснования, и не приводится никаких обсуждений для критической оценки надежности доказательств. Состояния хроматина — это динамические свойства клеток как in vivo, так и in vitro. На эти профили могут влиять физиологические условия и болезненное состояние. Насколько стабильны эти профили in vitro и насколько эти характеристики соответствуют их аналогам in vivo? Как эти профили различаются у разных людей по состоянию здоровья или болезни? А как насчет одноклеточной гетерогенности? Предположительно, эти факторы будут вносить шум в анализы, создавая ложноположительные и ложноотрицательные результаты.В принципе, все это нормально; у каждого подхода есть ограничения. Но удивительно мало внимания к этим вопросам ни при разработке исследования, ни при обсуждении результатов и анализа. Эти вопросы требуют внимательного, тщательного и критического рассмотрения.
Наши данные генерируются из трех реплик и / или доноров, и мы фокусируемся на регионах, которые доступны во всех репликах; действительно, мы отмечаем высокую согласованность между репликами. Следует отметить, что большинство различий в хроматине у разных людей являются количественными, а не качественными.Исследование, проведенное группой Регева (Gate et al., Nature Genetics 2018), показало, что открытые области хроматина в Т-клетках от> 100 человек были очень согласованными, но что 15% OCR варьировались количественно, что связано с общими генетическими вариациями. В сочетании с дополнительным ограничением настаивать на том, чтобы только SNP совпадали с открытым хроматином через ATAC-seq, наш подход, основанный на Capture C, позволяет избежать высокой степени ложных срабатываний; однако определить частоту ложноотрицательных результатов сложно из-за отсутствия справочных данных.Однако, учитывая статистическую значимость наших наблюдений, мы твердо верим, что наши данные должны использоваться в области генетики сложных заболеваний. Кроме того, мы гарантируем, что работаем только с клетками, происходящими от здоровых людей, учитывая, что мы сосредоточены на характеристике восприимчивости, присваиваемой этими локусами до начала заболевания, то есть мы исследуем генетическое строение до патогенеза заболевания, или, другими словами, « перед автокатастрофой ». Полученные здесь данные согласуются с наборами данных, которые мы описываем в недавних статьях, опубликованных в Nature Communications, для волчанки и минеральной плотности костей.Наконец, мы не проводили захват конформации хроматина в условиях одной клетки, подход, который все еще находится в зачаточном состоянии и имеет некоторые недостатки, включая важные этапы множественной амплификации, которые сами по себе могут привести к высокой степени ложноположительных результатов. Мы выделили эти моменты в Обсуждении.
10) Авторы должны обсудить, как модель может работать. Авторы справедливо обеспокоены неравновесием по сцеплению с упором на независимые доказательства (подраздел «Конвейер картирования вариантов к генам»).Но официально данные не представлены; если эти SNP независимы (без LD), каков аргумент в пользу того, что эти данные могут быть объединены, а не добавлены? Являются ли положительные результаты этого картирования от варианта к гену просто отражением аддитивных эффектов? Или они утверждают, что вовлечено SNP-взаимодействие (эпистаз)? Текст неясен по этим вопросам.
У нас менее ясны вопросы, поднятые рецензентом в этом отношении. На самом деле нас не беспокоит LD, скорее мы используем его для интеграции с данными ATAC-seq, чтобы составить короткий список предполагаемых причинных вариантов, которые в дальнейшем будут включены в окончательный список путем дальнейшей интеграции данных Capture C.Мы рассматриваем все независимые локусы BMI и WHR как независимые сигналы и делаем выводы как таковые. В нашем исследовании не рассматриваются и не наблюдаются какие-либо формы эпистаза.
11) Подход. Это важные вопросы для любого предложенного метода:
A) Авторы используют частотный подход. Возможно, они оправдывают этот подход в сравнении с байесовским подходом, которому хорошо соответствует логика их подхода.
Мы ценим это предложение. Мы обновили статистический анализ байесовской моделью, чтобы определить вероятность того, что наш метод с ограничениями превосходит любые ограничения.Мы отмечаем некоторые изменения в результатах, хотя ситуации, когда произошли изменения, привели к> 95% вероятности того, что наше биологическое ограничение превосходит никакие ограничения. И в целом изменения не влияют на нашу интерпретацию этих результатов.
B) Авторы должны быть в состоянии предоставить четкую базовую статистику — количество / процент совпадений, которые подтверждаются в целом, а затем, применяя их подход, по существу, они должны давать ложноположительные и ложноотрицательные показатели, т.е.е. сколько их подходов выдвигает, которые в конечном итоге оказываются отрицательными, и сколько они упускают из виду (предположительно, локусы с неэпигенетическими механизмами, очевидно, будут упущены). Очевидно, им необходимо четко указать, что применяется сопоставимый порог, чтобы читатели могли оценить относительную эффективность предлагаемых и стандартных методов. Что важно, сохраняют ли они уровень ошибок типа I с помощью своего нового метода и в чем их сила? К сожалению, в аннотации, результатах и обсуждении по этим вопросам нет ясности — это суть статьи.
Хотя мы сообщили значения чувствительности и специфичности в предыдущей версии рукописи, мы признаем, что ложноположительные и ложноотрицательные результаты не были включены напрямую. Мы сделали ссылку на общую тенденцию этих значений, учитывая их связь с чувствительностью и специфичностью, а также на общую картину этих показателей во всех анализах в разделе результатов, озаглавленном «Прогностическая сила отрицательного контроля не отличается от неограниченного набора», а также как в Обсуждении; однако мы признаем, что такая информация, касающаяся сохранения частоты ошибок типа I, не была включена.Чтобы учесть этот комментарий, мы сделали наш дополнительный файл данных более полным, который включает метрики классификации для каждого условия, а также теперь включает FPR и FNR. Впоследствии этот файл был переименован в Исходные данные 1. Кроме того, теперь мы включили более прямое обсуждение этих значений и их тенденций в раздел «Обсуждение».
https://doi.org/10.7554/eLife.62206.sa2Использование триаллельных SNP для определения происхождения …
Введение
Домашние яки ( Bos grunniens ) произрастают на Цинхай-Тибетском плато в Центральной Азии и ценятся во всем мире за их мясо, клетчатку, молоко, топливо, шерсть , транспортировка, защита от хищников и в качестве домашних животных 1 .Мировая популяция яков велика и разнообразна: более 14 миллионов домашних яков и 15 000 диких яков ( Bos mutus ) 2 . Напротив, североамериканская (NA) популяция яков небольшая и узкая, всего от 2 000 до 5 000 яков, все из которых являются домашними. Это стадо в основном возникло из нескольких десятков животных, завезенных из государственных и частных европейских зоологических парков в зоопарки Северной Америки на рубеже 20-го века. Отсутствие разнообразия источников также могло усугубить проблему.Например, Смитсоновский национальный зоопарк (SNZ) в Вашингтоне, округ Колумбия, импортировал своих первых яков в 1898 году из Лондонского зоологического общества, Англия 3 . В течение 23 лет закрытое стадо SNZ расширялось за счет размножения, а излишки животных отправлялись, пока зоопарк не получил единственного нового племенного быка яков из Парков Канады. Этот канадский бык произошел от яков, содержащихся в аббатстве Уобурн в Англии, примерно в 50 милях от первоначального источника в Лондонском зоопарке 4 . Помимо источников в зоологических парках, дополнительная зародышевая плазма яка не вводилась в стада АН из-за строгих федеральных нормативных ограничений, установленных на ввоз живых животных, эмбрионов и спермы.Таким образом, излишки яков с высокой степенью родства от ограниченных интродукций являются очевидными основателями нынешней популяции яков Северной Америки. Эта узкая генетическая база популяции NA-яков является постоянной проблемой для поддержания генетического разнообразия для защиты здоровья и жизнеспособности этих стад.
Задача сохранения генетического разнообразия яков NA усугубляется периодической гибридизацией с крупным рогатым скотом ( Bos taurus ). Проблемы, связанные с гибридизацией яков и крупного рогатого скота, возникли еще до их экспорта с Цинхай-Тибетского плато.Производство гибридов яка и крупного рогатого скота практикуется в течение 3000 лет, начиная с династии Инь 5,6 . Эти древние события гибридизации составляют 2–3% аллелей Bos taurus , присутствующих сегодня у домашних яков 7 .
Генетическое управление интрогрессией древних B. taurus у домашних яков обычно не вызывает беспокойства у заводчиков. Тем не менее, в небольшой популяции яков Северной Америки также зарегистрированы случаи интрогрессии крупного рогатого скота в начале 20-го века 8 , а также недокументированная интрогрессия в результате случайных усилий производителей по внедрению таких признаков крупного рогатого скота, как цвет шерсти и выход туши.В некоторых случаях существующие животные могут быть удалены только на пару поколений от предыдущего случая гибридизации крупного рогатого скота с яком F1. Таким образом, идентификация и документирование этих недавно гибридизированных животных (в пределах трех поколений) имеет важное значение для сохранения подлинной зародышевой плазмы Bos grunniens при производстве здоровых, генетически разнообразных животных.
Инструменты на основе SNP открывают новые возможности для более точного генетического управления определенным видом домашнего скота. Достижения в области обнаружения и тестирования SNP сделали повседневной проверку происхождения, идентификацию животных и отслеживание болезней крупного рогатого скота, овец, лошадей и свиней 9–12 .Ранее при тестировании происхождения яков использовались микросателлитные маркеры, полученные из Bos taurus (т.е. короткие тандемные нуклеотидные повторы) 13,14 . Однако общедоступные данные полных геномных последовательностей (WGS) китайцев и яков NA, вместе с разнесенной по гаплотипам сборкой F1 як-крупный рогатый скот, позволили идентифицировать информативные SNP NA яков in silico для потенциального развития маркеров родительских SNP. 15–17 . Конечная полезность набора родительских SNP может быть измерена их успехом в выполнении наиболее сложного сценария: «отслеживание одного родителя».Например, использование ДНК туши ягненка для идентификации его истинного отца в системе спаривания с несколькими отцами, но отсутствие ДНК матери для проведения анализа троицы 9 . Определение отца потомка требует генетического исключения всех других самцов, подвергшихся воздействию маток. Этот подход основан на том принципе, что настоящий отец должен иметь общий аллель с потомством на каждом участке тестирования 18 . Таким образом, когда потомство и потенциальный отец гомозиготны по разным аллелям в одном и том же сайте, потенциальный отец исключается из отцовства.Идеальный набор SNP для тестирования одного родителя имеет маркеры с частотой минорных аллелей (MAF) более 0,30, равномерно распределены по геному и имеют SNP с высококонсервативными фланкирующими последовательностями ДНК для эффективного и точного генотипирования 9 . При наличии таких SNP двуаллельного происхождения яка производители смогут получить доступ к коммерческому высокопроизводительному генотипированию SNP, использовать стратегии разведения на пастбищах с несколькими отцами, проверять родословные и устанавливать уникальную идентификационную информацию животных, которая может быть использована для отслеживания при необходимости.
Маркеры, подходящие для SNP двуаллельного происхождения яков, обычно не предоставляют информацию об интрогрессии B. taurus , что потребовало бы дополнительного набора двуаллельных SNP, тем самым увеличивая стоимость, ограничивая выбор платформ, подходящих для более низкой пропускной способности. Однако, основываясь на наших предыдущих межвидовых выравниваниях WGS 19 , мы выдвинули гипотезу, что некоторые сайты SNP двуаллельного происхождения у яка могут также выравниваться с почти мономорфным альтернативным аллелем в B.Телец . В этой гипотетической триаллельной системе SNP эволюционный источник всех трех аллелей может быть выведен путем оценки частот в выборках видов Bos и , рода, которые разошлись 5 миллионов лет назад. Эта идея согласуется с эволюцией рода Bos как комплекса генетически связанных видов с общими эволюционными траекториями 20 . Например, предковый аллель Bos будет обычным для большинства существующих видов Bos , включая яков (рис. 1A, нуклеотид «C»).Связанный с яком аллель, используемый для проверки отцовства, должен был возникнуть со времен появления последнего общего предка для видов Bos (рис. 1А, нуклеотид «Т»). Точно так же другой аллель, ассоциированный с B. taurus , мог бы возникнуть в те же временные рамки (рис. 1A, нуклеотид «A»). Эволюционное расстояние между B. taurus и B. grunniens может быть достаточным для того, чтобы каждый вид развил свои собственные отличные аллели в одних и тех же геномных сайтах, что позволяет идентифицировать новые триаллельные SNP по происхождению яков (tySNP) (рис. 1B).Если бы достаточное количество tySNP (например, один из 26 миллионов) могло быть идентифицировано с распределенным интервалом генома, высоким MAF у яков и высокой специфичностью у B. taurus , можно было бы разработать максимально информативный генетический тест с минимальный набор порядка 100 SNP. Для этого требуется всего лишь найти один высококачественный tySNP на 26 миллионов геномных позиций, и это было бы идеально для видов домашнего скота, селекционеры которых в настоящее время не имеют доступа к SNP с более высокой плотностью по всему геному и технологиям генотипирования.
Рис. 1. Гипотетическое распределение генотипов специализированного триллельного SNP для определения происхождения яков и оценки интрогрессии
B. taurus .Деревья видов Bos основаны на деревьях, опубликованных Wu et al . (Wu et al ., 2018. Nat Ecol Evol 2, 1139–45). ( A ) Дерево на основе полных аутосомных последовательностей с помощью метода рандомизированного осевого максимального правдоподобия (RAxML). ( B ) Дерево на основе полных аутосомных последовательностей с использованием метода алгоритма точного дерева видов (ASTRAL) с неперекрывающимся скользящим окном размером 100 Кбайт.* Дерево укоренено у основания ветки водяного буйвола ( Bubalus bubalus ). (Figure1_BosTrees.TIF)
Здесь мы описываем панель из 610 tySNP для использования в идентификации животных, тестировании отцовства и для оценки недавних событий интрогрессии B. taurus у NA яков. Маркеры были последовательно отфильтрованы на наличие: 1) идентичных гетерозиготных генотипов у NA и китайского эталонного яка, 2) идентичных гомозиготных генотипов у других нетауриновых видов Bos , 3) третьего высокочастотного аллеля в B.taurus , 4) консервативные фланкирующие последовательности у видов Bos и 5) уникальное место в геноме B. grunniens . Репрезентативный пробный набор из 87 tySNP был использован для оценки эффективности у NA яков с помощью платформы генотипирования MALDI-TOF. Основываясь на этих оценках, мы выполнили компьютерное моделирование, чтобы предсказать способность определять отцовство и недавнюю интрогрессию B. taurus у NA яков с увеличением числа tySNP. Наша конечная цель состояла в том, чтобы использовать минимальный набор tySNP для выполнения множества разнообразных генетических тестов.
Методы
Заявление об этике
Эта статья не содержит исследований, проведенных на животных. Первоначальные источники использованных архивных образцов ДНК были либо приобретены у компаний, которые собирали их для искусственного осеменения, а не для исследований (мясной скот), либо приобретены или переданы в дар от лиц, которые собирали их в частном порядке для своих целей, таких как пищевые (бизоны и яки), либо пожертвован Американскому центру исследования мясных животных (USMARC) частными лицами, которые собирали образцы в частном порядке для своих целей управления стадом (мясной скот и як, см. Благодарности ).Образцы ДНК бантенга (1 мкг каждый) были переданы за плату в соответствии с Соглашением о материалах исследования, которое было подписано 6 февраля 2007 года юридическими директорами зоопарка Сан-Диего и Управлением передачи технологий Министерства сельского хозяйства США, ARS. Образцы крови гаура были собраны в соответствии с протоколами IACUC зоопарка Омахи Генри Дурли 15 июня 1999 г. их директором по охране здоровья животных, доктором Дугласом Армстронгом, DVM, в присутствии соавтора, члена парламента Хитона. Животных оценивали на предмет межгосударственного перевода в другой зоопарк, для которого требовался забор венозной крови.Игла-бабочка использовалась для заполнения пробирок с кровью для необходимых исследований и сбора образцов ДНК.
Животные и WGS
Контрольной самкой домашнего яка NA, использованной для WGS и сопоставления с геномом B. taurus , была Queen Allante D171 (QA; Рисунок 2). QA умерла естественной смертью в январе 2010 года в возрасте примерно 30 лет и на момент своей смерти была самым старым из ныне живущих основателей популяции АН. Свежая шкура была собрана владельцем после забоя, заморожена при -20 ° C и отправлена в USMARC для экстракции ДНК и получения последовательностей генома.Примерно 20-кратный охват файлов FASTQ для обеспечения качества был получен из инвентарного номера BioProject PRJNA325061 (BioSample SAMN05558793, SRX2026482, SRX2026483 и SRX2026485 — SRX2026494). Подробности подготовки библиотеки и секвенирования описаны Heaton et al . 21 .
Китайская эталонная самка домашнего яка, использованная для выравнивания с геномом B. taurus , была Qh2 15 . Приблизительно 60-кратный охват файлов FASTQ был получен из регистрационного номера BioProject PRJNA74739 (BioSample SAMN00744358, SRX103173-SRX103192).Первоначально использовался набор данных об этом животном, поскольку это был единственный набор данных о яках с достаточным охватом (> 10 раз) как для обнаружения вариантов, так и для точного генотипирования. Впоследствии дополнительные данные о яках были доступны из инвентарного номера BioProject PRJNA285834, включая B. grunniens : DYS74, SAMN03766772, SRX1056027; DYS77, SAMN03766773, SRX1056028; DYX12, SAMN03766774, SRX1056029; DYX11, SAMN03766775, SRX1056030; DYY31, SAMN03766776, SRX1056031 и B. mutus : WYX01, SAMN03766777, SRX1056032; WYX02, SAMN03766778, SRX1056033; WYX03, SAMN03766779, SRX1056034; WYX04, SAMN03766780, SRX1056035; и WYX05, SAMN03766782, SRX1056036 22 .
WGS и объединение як-крупный рогатый скот F1 с гаплотипами «Esperanza», теленка самки яка NA и быка говядины Highland 17 , также использовались для оценки точности tySNP (ARS_UNL_BGru_maternal_1.0_p), BioProject номера доступа PRJNA551500 и PRJNA552915 (BioSample SAMN12153487, SRR12094761). Кроме того, для оценки частот аллелей в подмножестве 87 tySNP были использованы исследовательские и коммерческие данные генотипа из 170 образцов NA яков, полученных из 36 источников.
В комиссию по мясному скоту вошли 96 неродственных особей из 19 популярных U.S. beef Breeds (USMARC Beef Diversity Panel, версия 2.9 [MBCDPv2.9]) 21 . Родословные были получены от ведущих поставщиков спермы крупного рогатого скота США и ассоциаций пород и проанализированы для выявления неродственных особей для включения. На основе количества зарегистрированного потомства породы, по оценкам, представляют более 99% зародышевой плазмы, используемой в мясном скотоводстве США, содержат более 187 неразделенных гаплоидных геномов и позволяют с 95% вероятностью обнаружить любой аллель с частота больше 0.016 23 . Породы в MBCDPv2.9 были (в порядке убывания зарегистрированного потомства около 2000 г.): ангус (n = 6), герефорд (n = 6), шароле (n = 6), симментальский (n = 6), красный ангус (n = 6). = 6), Лимузен (n = 6), Гельбви (n = 6), Брангус (n = 5), Beefmaster (n = 5), Салерс (n = 5), Шортхорн (n = 5), Мэн-Анжу ( n = 5), Brahman (n = 6), Chianina (n = 4), Texas Longhorn (n = 4), Santa Gertrudis (n = 4), Braunvieh (n = 4), Tarentaise (n = 4) и Корриенте (n = 4). Средний охват генома в файлах FASTQ для этих 96 мясных быков был примерно 14-кратным и доступен в NCBI SRA со ссылками на регистрационный номер BioProject PRJNA324822 21 .
Другие использованные виды Bos включали примерно 10-14-кратное покрытие WGS от гаура (n = 2), бантенга (n = 2) и бизона (n = 1) и были получены из инвентарного номера BioProject PRJNA325061. в том числе B. gaurus : 199
1, SAMN05558794, SRX2026439-SRX2026446, SRX2026451, SRX2026462, SRX2026473, SRX2026484, SRX2026495 — SRX2026498; 199
2, SAMN05558795, SRX2026447 — SRX2026450, SRX2026452 — SRX2026461, SRX2026463 — SRX2026464; B. javanicus : 200710001, SAMN05558796, SRX2026465 — SRX2026468; 200710002, SAMN05558797, SRX2026469 — SRX2026472; и Б.Бизон : 1991, SAMN05558798, SRX2026474-SRX2026481 21 .
Картирование считывания и обнаружение вариантов
Все данные о последовательностях, использованные в этом исследовании, были обрезаны по качеству с помощью TrimGalore (версия 0.5.0) и сопоставлены со сборкой Bos taurus , UMD3.1 24 , с помощью выравнивателя Burrows Wheeler ( версия 0.6.1) 25 модуль aln . Затем файл в формате sam был преобразован в формат bam и отсортирован с помощью samtools (версия 0.1.18) 26 .Samtools также использовался для маркировки дубликатов ПЦР с помощью функции «markdup». Модуль IndelRealigner из набора инструментов анализа генома 27 (GATK версия 1.5-32-g2761da9) Института Броуда использовался для обеспечения того, чтобы любые вставки или удаления были согласованы друг с другом. Обнаружение вариантов и генотипирование для картированных наборов данных WGS для животных яков, гаур, бантенгов и бизонов (n = 7) были выполнены с использованием модуля GATK UnifiedGenotyper (версия 3.4-46-gbc02625), запущенного с genotyping_mode = DISCOVERY. Файл VCF был отфильтрован с помощью специальной программы (ParseYakCoarse.java) 28 для идентификации tySNP, где животные гаура, бантенга и бизона (n = 5) были гомозиготными по одному и тому же предполагаемому предковому аллелю Bos (например, рис. 1A, «C / C»), и оба эталонных яка были гетерозиготными. для предкового аллеля Bos и аллеля, ассоциированного с яком (например, рис. 1A, «C / T»). Ассоциированный с яком аллель не присутствовал в сборке бычьего UMD3.1 или трех других видов Bos , а предковый аллель отличался от аллеля, присутствующего в UMD3.1 сборка
Скрининг третьего распространенного аллеля у мясного скота
Файл VCF, содержащий эти отфильтрованные выше SNP, затем был использован в качестве аргумента —alleles для унифицированного генотипа с genotyping_mode = GENOTYPE_GIVEN_ALLELES и 96 файлов BAM из разнообразия крупного рогатого скота панель как входные файлы BAM. Опция «—intervals» также использовалась с файлом .bed, в котором была запись, определяющая локус 2001 базисов с центром на каждом из SNP. Это ограничило объем генотипирования только недавно отфильтрованными SNP и позволило получить генотипы для всех 96 голов крупного рогатого скота по этим SNP.Специальная программа (ParseYakFine.java) 28 была написана для фильтрации записи VCF, пропускающей только те записи, в которых 183 или более из 192 возможных аллелей крупного рогатого скота (т. Е.> 95%) были специфичными для крупного рогатого скота UMD3.1 крупного рогатого скота. референсный аллель (например, рис. 1A, «A / A»). Более половины записей в результирующем файле VCF имели значение FILTER LowQual, поскольку в этих записях ни у одного животного не было обнаружено нереференсных аллелей. Чтобы использовать полученный VCF в качестве входных данных для следующего шага, это значение ФИЛЬТРА было отредактировано с помощью текстового редактора emacs, а «LowQual» было заменено на «.». Отредактированный файл был использован в качестве исходных данных для генотипирования WGS.
Генотипирование tySNP у диких и домашних китайских яков
VCF-файл, содержащий кандидаты tySNP, использовался в качестве аргумента —alleles в программе Unified Genotyper с genotyping_mode = GENOTYPE_GIVEN_ALLELES, и были генотипированы файлы bam, созданные для пяти диких и четырех домашних китайских яков. . Опция «—intervals» также использовалась с файлом .bed, в котором была запись, определяющая локус 2001 оснований, центрированных на каждом из полиморфизмов, чтобы ограничить область генотипирования теми локусами, которые имеют отношение к данному исследованию.Специальная программа (ParseVCF_OtherYak.java) 28 была написана для подсчета количества наследственных, ассоциированных с яками и B. taurus -ассоциированных аллелей, присутствующих у каждого яка.
Извлечение соседних консервативных последовательностей, идентификация соседних SNP и маскирование
Положение хромосомы B. taurus UMD3.1 и аллели для маркеров-кандидатов использовались с пользовательской программой (ParseYakFinal.java) 28 для извлечения 100 оснований фланкирующей последовательности как на 3 ‘, так и на 5’ позиции маркера.Файлы bam для пяти диких и четырех домашних китайских яков и панель 96 разнообразия крупного рогатого скота 21 были использованы для идентификации соседних SNP, которые могут нарушать образование гетеродуплекса с помощью олигонуклеотидов, используемых в анализах генотипирования на различных платформах генотипирования. GATK UnifiedGenotyper был запущен с genotyping_mode = DISCOVERY, а значение —intervals было установлено как файл .bed, содержащий записи для всех локусов, указанных с 2001 базами, центрированными на каждом маркере. Варианты крупного рогатого скота, обнаруженные в последовательностях 100 п.н., соседних с tySNP с обеих сторон, были заменены на «N», если их частоты аллелей были больше или равны 5% (т.е.е., 9 или более из 192 возможных аллелей). Эти вариантные флаги могут использоваться программным обеспечением для разработки анализа генотипа для перенаправления размещения олигонуклеотидов на более консервативные фланкирующие последовательности. Точно так же варианты yak, фланкирующие tySNP на любой частоте (по сравнению со сборкой B. taurus UMD3.1), также были заменены на N по той же причине.
Выравнивание tySNP с геномом яка для определения положения хромосомы и назначения кластера
Последним шагом в фильтрации кандидатов tySNP было выравнивание 200 п.н. фланкирующей последовательности с разрешенной гаплотипом сборкой яка F1 (ARS_UNL_BGru_maternal_1.0_p). Это определило хромосому яка и положение tySNP. Маркеры исключались из группы, если они не отображались в сборке ровно один раз. Остальные tySNP были вручную отнесены к маркерным «ячейкам» на основе их положения в хромосомах. Кластерные маркеры (например, менее 1 МБ) обычно группировались в одном бункере, чтобы дать возможность выбора программного обеспечения для разработки анализа SNP для наиболее подходящей мишени в регионе. По возможности, желаемый интервал между ячейками составлял 5 Мб. При идеальном интервале 5 МБ в аутосомном геноме яка размером 2479 МБ будет около 496 интервалов плюс 29 для конечных интервалов.
Матричная лазерная десорбция / ионизационная времяпролетная масс-спектрометрия (MALDI-TOF MS) Дизайн анализа для подмножества 87 tySNP
Перед разработкой анализа пороговые значения для частоты вызовов (общее количество полученных генотипов / общее количество возможных генотипов) ) и точность (правильные генотипы / все полученные генотипы) были установлены на 97% и 99% соответственно. Для целей исключения отцовства показатель отсечения 97% означает, что минимум 94% tySNP будут иметь генотип для каждого из двух животных при попарном сравнении.Хотя эти пороговые значения относительно высоки, мы считаем их «золотым стандартом» в тестировании отцовства на основе SNP и находятся в пределах возможностей сегодняшней технологии тестирования ДНК. Наборы SNP по происхождению, соответствующие этим стандартам, существенно повышают эффективность тестирования 9 . Разработка анализа и генотипирование выполнялись в Neogen Genomics (Линкольн, Небраска, США) с платформой MassARRAY и химическим анализом iPLEX GOLD в соответствии с инструкциями производителя (Agena Bioscience, Сан-Диего, Калифорния, США).Мультиплексные анализы были разработаны с использованием программного обеспечения для разработки анализов производителя и предварительного набора из 139 бункеров с 518 tySNP, выровненных со сборкой B. taurus UMD3.1. Параметры программного обеспечения были установлены для максимум 48 анализов на одно сплетение и для выбора одного маркера tySNP, доступного в любой ячейке. Концентрации зонда-удлинителя были отрегулированы эмпирически для оптимизации сигнала по всему масс-спектру. Три мультиплексных анализа были выполнены на ДНК контрольных яков NA (QA) и других NA-яков. Конкретные анализы SNP, которые давали низкую частоту вызовов или высокую частоту ошибок, подвергались цензуре на основе наборов данных.Для настоящего отчета данные генотипа использовались для любого из этих 139 объединенных SNP, которые соответствовали критериям анализа, указанным выше, и имели уникальную позицию на картировании в сборке яков F1 с разрешенным гаплотипом.
Оценка вероятности идентичности (P
I ) и вероятности исключения (P E ) с tySNPP I является оценкой вероятности совпадения генотипов у двух животных. Допущения, использованные для анализа tySNP, включали распределение генотипов по Харди-Вайнбергу (HW), пренебрежимо малую частоту B.taurus -ассоциированный аллель, и что средний MAF для аллелей отцовства яков у NA yak является репрезентативным. Вкратце, P I для локуса A с аллелями SNP A 1 и A 2 представлял собой сумму квадратов частот трех генотипов: P I = (χ 11 ) 2 + ( χ 12 ) 2 + (χ 22 ) 2 , где χ 11 , χ 12 и χ 22 были относительными частотами генотипов A 1 A 1 , A 1 A 2 и A 2 A 2 , соответственно 29 .Комбинированный P I для нескольких маркеров SNP был продуктом P I для каждого отдельного маркера. Основное предположение заключалось в том, что расстояние между маркерами было достаточным для мейотической рекомбинации, чтобы вызвать случайную ассоциацию аллелей друг с другом. Однако с увеличением плотности родительских SNP обоснованность этого предположения снижается. Таким образом, объединенный P I для более чем одного родительского SNP на хромосому является недооценкой вероятности случайного совпадения между случайными животными из популяции из-за неравновесия по сцеплению между SNP на одной и той же хромосоме.Кроме того, для целей оценки P I и P E мы предположили, что частота аллеля, ассоциированного с B. taurus (A 3 ), была незначительной.
P E — вероятность того, что случайное животное будет исключено из отцовства. P E также является наименее сложным методом анализа отцовства и оценивает долю потенциальных взрослых, исключенных из отцовства. В этом отчете во всех оценках P E строго использовалась информация только о генотипе одного из родителей i.е., самый сложный сценарий. Таким образом, исключение было основано только на частоте противоположных гомозиготных генотипов SNP у потомства и предполагаемого родителя, как описано ранее 9 . Вкратце, вероятность противостояния гомозигот (P OH ) между случайным потомком и случайным подходящим взрослым в локусе A SNP с аллелями A 1 и A 2 рассчитывалась следующим образом: P OH = (χ 11 потомство) (χ 22 взрослый) + (χ 22 потомство) (χ 11 взрослый), где χ 11 и χ 22 были относительными частотами генотипов A 1 A 1 и A 2 A 2 , соответственно, для групп взрослых или потомков.Предполагалось, что частоты гомозиготных генотипов SNP одинаковы в породной группе независимо от возраста. Таким образом, для одного двуаллельного SNP P E = P OH = 2 (χ 11 ) (χ 22 ), когда один из генотипов родителя недоступен. Это представляет собой долю подходящих взрослых, которые были бы исключены из отцовства в одном локусе, усредненную по всем сравнениям между потомством и взрослыми. Без использования информации о генотипе другого родителя комбинированный P E для нескольких SNP был следующим: P E (SNPn) = P E (SNP1) + R 1 P E (SNP2) + R 2 P E (SNP3) … + R n-1 P E (SNPn) , где P представляет собой долю отвечающих критериям взрослых, исключенных первым SNP, а 1 R — оставшаяся часть не исключенных. взрослые люди.R 2 — R n-1 — это оставшиеся доли не исключенных взрослых после каждого раунда последующего тестирования с n родительскими SNP. Таким образом, для 29 SNP по происхождению (по одному на каждой аутосоме) комбинированный P E для неродственных родителей определяется следующим образом: P E (29) = P E (1) + R 1 P E ( 2) + R 2 P E (3) … + R 28 P E (29) . Как и в случае с комбинированным P I , комбинированный P E для более чем одного родительского SNP на аутосому является недооценкой вероятности того, что случайный предполагаемый родитель будет исключен из отцовства из-за неравновесия по сцеплению между SNP на одном и том же хромосома.Для родственных родителей P E для каждого SNP умножали на коэффициент родства ( r ), где r = 0,125, 0,250 или 0,500 [34]. Таким образом, P E (29) для родственных родителей = ( r P E (1) + r R 1 P E (2) + r R 2 P E (3) … + r R 29 P E (29) ).
Выявление исключений в системе трехаллельных SNP
Триаллельные SNP могут приводить к исключению по происхождению способами, которых не делают двуаллельные, из-за присутствия B.taurus -ассоциированные аллели у некоторых людей. Рассмотрим три аллеля tySNP: предковый аллель Bos (A 1 ), родительский аллель яка (A 2 ) и аллель, ассоциированный с B. taurus (A 3 ). Исключение происходит, когда у теленка и взрослой особи нет общих аллелей. Таким образом, при оценке сценариев «одного родителя», в которых генотипы матери недоступны, анализ не ограничивается только противоположными гомозиготными генотипами.Гетерозиготные сайты могут стать информативными, если теленок или взрослый особь обладают копией аллеля, ассоциированного с B. taurus . Двумя распространенными примерами этих конфигураций генотипа являются: теленок — A 1 / A 1 , а взрослый — генотип A 2 / A 3 , или когда теленок — A 1 / A 3 . а взрослый — A 2 / A 2 . В обоих примерах взрослый исключен из отцовства. Аналогичным образом, при оценке сценариев «с двумя родителями», где генотип матери на некоторых участках может использоваться для определения аллеля производителя у теленка, гетерозиготный участок в матери и теленке может стать информативным, если тот или другой обладает копией модель B.taurus -ассоциированный аллель. Например, если теленок — A 1 / A 2 , а мать — A 2 / A 3 , аллель производителя теленка — A 1 и исключает всех взрослых, не несущих A. 1 аллель. Хотя исключения по происхождению, вызванные аллелями, ассоциированными с B. taurus , встречаются только в нескольких процентах сравнений генотипов яков NA, их важно учитывать при обработке тестов на отцовство в коммерческих условиях.
Моделирование для оценки силы исключения с помощью tySNP при наличии ошибки генотипирования
Триаллельный SNP имеет повышенный потенциал для исключения случайных взрослых из отцовства по сравнению с двуаллельным SNP. Однако чрезвычайно низкая частота ассоциированного с B. taurus аллеля у яков приводит к незначительному вкладу в исключение яков по происхождению. Следовательно, третий аллель был проигнорирован для целей этого моделирования. Один миллион случайных пар потомство / взрослый был смоделирован с частотами аллеля tySNP, выведенными из генотипов 170 яков NA с допущениями HW.Пара потомство / взрослый приблизительно соответствует сценарию тестирования одного родителя, поскольку генотипы другого родителя не использовались для определения фаз гетерозиготных сайтов потомства. Исключение из отцовства подсчитывали для каждого сайта tySNP, где потомство и взрослый были гомозиготными по разным аллелям. Распределение частот исключений для данного набора tySNP определяли путем суммирования эксклюзивных сайтов для каждой пары потомство / взрослый по всем парам. Второе моделирование было выполнено для проверки эффекта «выпадения аллеля», хорошо известного источника ошибки генотипирования, и систематической ошибки, которую мы чаще всего ожидаем от высококачественной выборки в этой панели SNP.Это происходит, когда другие геномные SNP присутствуют в сайтах связывания для трех олигонуклеотидов, используемых в анализе MALDI-TOF. Эти SNP могут нарушать образование гетеродуплекса в любом из трех необходимых праймеров для анализа и вызывать отсутствие связанного целевого аллеля в генотипированных аллелях. В этом моделировании был смоделирован производитель яка путем выбора аллелей для каждого tySNP на основе частот аллелей, выведенных из генотипов 170 NA Yak с допущениями HW. Для каждого tySNP один из аллелей производителя yak был назначен потомству, а второй аллель был выбран случайным образом на основе частот аллелей для соответствующего маркера.Затем к каждому генотипу применялась фиксированная частота ошибок генотипа. Для тех генотипов, которые были выбраны как ошибочные, один из двух аллелей не учитывался в вызове, и событие записывалось, если было введено исключение артефактов.
Моделирование для оценки способности tySNP выявлять недавние события гибридизации F1
Событие гибридизации яка / крупного рогатого скота F1 легко обнаруживается с помощью tySNP, поскольку потомство будет иметь аллель, ассоциированный с B. taurus на каждом участке (рис. 3) .Однако сколько может произойти обратных скрещиваний с яком, прежде чем аллели крупного рогатого скота больше не будут надежно обнаружены? Вероятность передачи аллеля крупного рогатого скота составляет 0,5 для каждого поколения после события F1. Таким образом, можно ожидать, что потомство будет терять в среднем половину аллелей, ассоциированных с B. taurus , в каждом последующем поколении (рис. 3). Это приведет к примерно 50%, 25% и 12,5% в первом, втором и третьем поколениях, соответственно, после события F1. Моделирование было выполнено, где гибрид яка / крупного рогатого скота F1 имел один аллель крупного рогатого скота и либо предковый, либо связанный с яком аллель Bos в каждом из положений tySNP.От этого смоделированного потомства F1 было также смоделировано скрещивание с яком, у которого не было аллелей крупного рогатого скота ни по одному из tySNP. Это было достигнуто путем случайного удаления одного из двух аллелей F1 на каждом участке и добавления второго аллеля, не относящегося к крупному рогатому скоту, который должен был быть внесен в результате обратного скрещивания. Начиная с генотипов, полученных в результате этого имитационного спаривания, новый набор генотипов был сгенерирован с использованием того же процесса для дополнительных пяти поколений. Затем было построено распределение оставшихся аллелей крупного рогатого скота на поколение.
Рис. 3. Выявление аллелей крупного рогатого скота в результате гибридизации F1 с последующим обратным скрещиванием яков.
Сокращения: P1 — родительское поколение; F1, гибридное поколение; B — обратное скрещивание поколений; нет данных, не применимо. (Figure3_CattleIntrogressionChart3.TIF)
Результаты
Идентификация кандидатных tySNP
Согласование эталонных последовательностей генома яка QA и Qh2 с эталонной сборкой генома B. taurus позволило идентифицировать 3133 кандидатных tySNP, которые были: 1) гетерозиготными 2) в обоих яках. гомозиготный по гаурам, бантенгу и бизону, и 3) имел третий B.taurus -ассоциированный аллель. Последующая фильтрация менее частых аллелей крупного рогатого скота (т. Е. Менее 0,95) уменьшила набор tySNP до 1023. Выравнивание фланкирующих последовательностей 1023 кандидатных tySNP с B. grunniens с разрешенной по гаплотипом сборке генома яка идентифицировало 612 tySNP с уникальными положения координат хромосомы. Два дополнительных SNP-кандидата были удалены как мономорфные после того, как все tySNP были генотипированы у 170 NA яков в финальном раунде с обновленной статистикой.Остальные 610 tySNP были сгруппированы в 441 региональный кластер (т. Е. Бункеры) со средним расстоянием между бинами 5,26 Мб (таблица S1) 30 . Из 610 tySNP 87 совпали с неопубликованным набором tySNP, для которых мы ранее разработали анализы генотипирования MALDI-TOF MS (таблица S2, см. методы ) 31 . Эти 87 tySNP были подмножеством маркеров, идентифицированных с помощью того же процесса, за исключением выравнивания с разрешенным гаплотипом генома яка. Их предыдущий выбор предшествовал появлению этой новой эталонной сборки яков.Таким образом, их уникальное выравнивание и положение генома яка относительно эталонной сборки яка ранее не были подтверждены. Положения 87 tySNP вместе с другими 523 tySNP показаны на гаплотипически разрешенной сборке генома яка (Рисунок 4). Вместе все 610 tySNP, их бины и информация о последовательностях подходят для ввода в программное обеспечение для разработки анализа SNP для различных платформ генотипирования.
Рис. 4. Хромосомное расположение tySNP.
Позиции 610 tySNP показаны выровненными по ARS_UNL_BGru_maternal_1.0_p як в сборе. Желтыми точками показаны 87 tySNP, преобразованные в анализы и используемые в NA yak для оценки частот и полезности аллелей. (Figure4_610tySNPs_442bins4.TIF)
Частоты аллелей 87 выбранных tySNP
Генотипы из анализов MALDI-TOF MS для 87 tySNP были оценены в 170 NA yak из 36 источников для оценки частот аллелей родительских аллелей и аллелей крупного рогатого скота ( Таблица S5) 32 . Общая частота генотипирования SNP (то есть «частота звонков») была равна 0.9952 для 87 тыСНП у 170 животных. Предковый аллель Bos был основным аллелем для большинства tySNP у NA яков (67%). Среднее значение MAF для аллеля происхождения яка составляло 0,296, причем 53% из них составляли пороговое значение 0,30 (рис. 5А). Общая частота аллелей, ассоциированных с B. taurus , была очень низкой (0,0043), при этом более половины яков NA имели ноль из 174 возможных аллелей крупного рогатого скота среди 87 протестированных участков (рис. 5В). Частоты аллелей, ассоциированных с B. taurus и , для шести из 87 tySNP были выше, чем у остальной части группы, хотя и меньше 0.1 в целом. (Рисунок 5C).
Рис. 5. Распределение частот аллелей 87 tySNP на основе генотипов 170 яков NA.
( A ) Як по происхождению SNPs MAFs. ( B ) B. taurus -ассоциированные аллели у животных. Панель : C , вариация частот B. taurus -ассоциированных аллелей среди различных tySNP. (Figure5_87_SNPs_MAFCatt4.TIF)
Идентификация животных и исключение по происхождению с 87 SNP
Использование среднего MAF для 87 tySNP (0.296) и предположения HW, P I для одного SNP составлял 0,427, что означает, что примерно 43% NA яков, как ожидается, будут иметь идентичные генотипы при среднем tySNP. Распространение этого на 29 tySNP, равномерно распределенных на 29 аутосомах, дало комбинированное теоретическое значение P I , равное 1,92 × 10 -11 . Однако выбор лучших tySNP с высоким MAF дал немного лучший результат только с 28 аутосомами: P I = 5,81 × 10 -12 (рис. 6A, таблица S2) 31 .Для определения производителя теленка без генетической информации о матери (т. Е. Тестирования однопородного отцовства) теоретическое значение P E для одного SNP составило 0,087 при тех же предположениях MAF и HW. Таким образом, примерно 9% кандидатов в быки могут быть исключены с помощью одного tySNP. Чтобы исключить группу из 30 быков-кандидатов на уровне достоверности 99%, потребуется 87 tySNP, все с одинаковым P E (Рисунок 6B). Было на 19 быков меньше исключено с уровнем достоверности 99% при использовании фактического P E , рассчитанного на основе генотипов NA для всех 87 tySNP, вероятно, из-за связи между tySNP и низким MAF в некоторых маркерах.При добавлении генотипов матери сила исключения увеличивается примерно вдвое благодаря способности фазировать гетерозиготные аллели теленка.
Рис. 6. P
I и P E оценки для tySNP у яков NA.( A ) Вероятность случайного совпадения генотипа с tySNP. ( B ) Количество tySNP, необходимое для исключения 99% быков-кандидатов из отцовства при отсутствии информации о генотипе матери. (Рисунок 6_PI_PE_7.TIF)
Распределение аллелей 610 tySNP, генотипированных
in silico с WGS, и сравнение с выбранными 87 SNPВнутренние свойства и распределение аллелей 610 tySNP были дополнительно оценены in silico с WGS от яков, Bos виды, мясной скот и гибридное трио як-крупный рогатый скот F1. По замыслу, два эталонных яка были гетерозиготными во всех 610 сайтах, имея ровно один предковый аллель Bos и один ассоциированный с яком аллель (Таблица 1).Исключая монозиготных близнецов, это единственные два яка, которые, как ожидается, будут иметь все 610 идентичных гетерозиготных генотипов, поскольку скрининг маркеров tySNP был нацелен на них. Также по дизайну частота предкового аллеля Bos была зафиксирована у гаура, бантенга и бизона из-за выбора этого свойства при фильтрации. Заметным исключением был единственный tySNP у бизонов, который был гетерозиготным по 4-му аллелю в одном сайте (A / T, ARS1.2-UCD chr12: 83562937, таблица S2) 31 . Неожиданный результат генотипа был обнаружен в одном из пяти наборов данных по домашнему китайскому яку (DYY31, SAMN03766776), который содержал более 98% B.taurus -ассоциированные аллели. Дополнительный анализ, проведенный на его нанесенном на карту наборе данных WGS (таблица S6) 33 подтвердил, что это набор данных B. taurus , и он был исключен из последующих анализов.
Таблица 1. Частоты аллелей 610 tySNP в наборах данных WGS.
| Аллели | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Животное или группа | No. — связанный | ||||||||||||||
| NA исх.як (QA) | 1 | 1.0000 | 0.5000 | 0.5000 | — | ||||||||||
| Китайский исх. Як (Qh2) | 1 | 1,0000 | 0,5000 | 0,5000 | — | ||||||||||
| Гаур | 2 | 1,0000 | 1,0000 | – | 101,0000 | — | — | ||||||||
| Зубр | 1 | 1.0000 | 1,0000 | — | — | ||||||||||
| Китайские дикие яки | 5 | 0,9790 | 0,6237 | 0,3721 | 0,0042 | ||||||||||
| 0,0042 | |||||||||||||||
| 0,0076 | |||||||||||||||
| Мясной рогатый скот | 96 | 0,9997 | 0,0104 | 0,0028 | 0,9865 | ||||||||||
| Хайленд бык (отец) | 1 | 1 | 1 99984 | 0,0016 | — | 0,9984 | |||||||||
| NA як (плотина) | 1 | 1.0000 | 0,5459 | 0,4443 | 0,0082 | ||||||||||
| 915 | 0,2738 | 0,2213 | 0,5049 | ||||||||||||
Остальные наборы данных о китайских яках были проанализированы на предмет предковых аллелей и содержания аллелей крупного рогатого скота. Средняя частота предковых аллелей была немного выше у диких яков (0.624) по сравнению с домашними яками (0,608), тогда как частоты аллелей, ассоциированных с B. taurus , у этих яков были ниже (0,0042 и 0,0076, соответственно, таблица 1). Частоты этих B. taurus -ассоциированных аллелей у китайских яков сравнивают с 0,0043, оцененными для 87 tySNP, генотипированных у NA-яков. У мясного скота частота предкового аллеля Bos составляла всего 0,010 из-за отбора в процессе фильтрации аллелей, ассоциированных с B. taurus . Частота Б.taurus -ассоциированных аллелей в 610 tySNP у мясного скота было 0,9865. Эти B. taurus -ассоциированные частоты аллелей соответствовали генотипам WGS из трио гибридного семейства F1 як-крупный рогатый скот: 0,9984 для хайлендского производителя, 0,0082 для самки яков NA и 0,5049 для теленка F1. Таким образом, внутренние свойства и распределение аллелей 610 tySNP, генотипированных in silico с помощью WGS, соответствовали тем, которые были получены в мультиплексных анализах MALDI-TOF MS для 87 tySNP в группе из 170 яков NA из 36 источников.
Моделирование исключения происхождения с помощью больших наборов tySNP
Исходя из внутренних свойств 87 tySNP, указанных выше, разумно экстраполировать их производительность в сценарии с большими наборами tySNP. Эффективное исключение отцовства зависит от ряда факторов, включая: MAF, количество маркеров и частоту ошибок генотипирования. Это также требует, чтобы большинство случайных не-родителей имело значительно больше исключений (т. Е. Противоположных гомозиготных генотипов), чем ошибочные исключения у истинного родителя из-за ошибок генотипирования.С текущим набором из 87 tySNP и средним MAF 0,296, большинство случайных пар теленок-взрослый имели от 5 до 10 исключительных генотипов (т. Е. Противоположных гомозиготных генотипов), в то время как большинство пар детеныш-родитель имеют только 0-2 ложных исключительных генотипа при 1%. частота ошибок генотипирования (рис. 7А). При 5% вероятности ошибок генотипирования ожидается, что примерно 0,05 истинных родителей будут иметь три ложных исключения генотипа и попадут в перекрытие с 0,03 правильных исключений у не-родителей. Это означает, что 0,0015 пар детеныш-взрослый будет трудно исключить из отцовства из-за совпадения ложных исключений у истинных родителей и правильных, но немногочисленных исключений у не-родителей.Это перекрытие можно улучшить, увеличив количество похожих tySNP, используя маркеры с более высокими MAF, или уменьшив частоту ошибок генотипирования (рис. 7B и 7C). Кроме того, если генотипы матери доступны для одних и тех же tySNP, мощность существенно увеличивается вдвое.
Рис. 7. Смоделированная мощность исключения отцовства с расширенными наборами tySNP, ошибками генотипирования и без генотипов матери.
Моделирование проводилось, как описано в Методах. Красные и розовые стрелки указывают на неоднозначное совпадение между правильными исключениями у лиц, не являющихся родителями, и ложными исключениями у настоящих родителей при 5% и 1% вероятности ошибок генотипирования, соответственно.(Figure7_ParentageSim4.TIF)
Мощность для обнаружения недавней интрогрессии крупного рогатого скота
Способность обнаруживать недавнюю интрогрессию крупного рогатого скота после гибридизации F1 и последующего обратного скрещивания яков моделировалась с помощью наборов tySNP. Приблизительно половина аллелей крупного рогатого скота гибрида F1 теряется в последующих поколениях обратного скрещивания и, таким образом, соответствует модели биномиального распределения. При наличии 87 tySNP у животного в третьем поколении обратного скрещивания (т. Е. 15/16 яка) можно ожидать обнаружения 10 B. taurus -ассоциированных аллелей (рис. 8A).Это сравнивается с полнокровным NA-яком, у которого в среднем было менее 1 аллеля, ассоциированного с B. taurus на животное (рис. 5B). Удвоение tySNP увеличивает уровень обнаружения для другого поколения обратного скрещивания (рис. 8B и 8C). Виды и формы этих кривых были бы сложнее и многочисленнее, если бы в недавней родословной одного яка было более одного скрещивания крупного рогатого скота / яков. В любом случае, NA-як с менее чем 3 из 87 B. taurus -связанных аллелей вряд ли будет иметь гибрид F1 в качестве родителя, прародителя или прадедушки.
Рис. 8. Имитация обнаружения интрогрессии крупного рогатого скота с помощью tySNP.
( A ), ( B ) и ( C ) Результаты моделирования с 87, 174 и 350 tySNP соответственно. Каждый пик представляет одно поколение обратного скрещивания, удаленное из события гибридизации F1. (Figure8_tySNP_BackCrossSim7.TIF)
Обсуждение
Наша цель состояла в том, чтобы идентифицировать набор новых триаллельных SNP. Мы идентифицировали новый набор из 610 tySNP, где каждый маркер имеет два аллеля для тестирования отцовства NA-яков и третий аллель для идентификации недавней интрогрессии крупного рогатого скота.Данные по дизайну анализа для этих маркеров представлены в таблице S1 30 . Помимо частот аллелей, маркеры были тщательно отобраны для уменьшения отрицательных атрибутов, таких как отступы, повторяющиеся последовательности и фланкирующие SNP, чтобы повысить их эффективность в существующих и будущих технологиях генотипирования. TySNP были распределены по геному в 441 кластере (бункере) со средним интервалом 5 Мб на основании недавно завершенной сборки генома NA yak 17 . Подмножество 87 tySNP было разработано для платформы MALDI-TOF MS и их способности: 1) предоставлять каждому животному уникальный генетический идентификатор, 2) исключать случайных взрослых особей для определения происхождения и 3) идентифицировать животных с событиями гибридизации F1 в их недавнее происхождение.При выборе лучших tySNP на каждой доступной аутосоме только 28 tySNP из 87 были необходимы для однозначной идентификации каждого NA-яка (по оценкам в совокупности P I = 5,81 × 10 -12 ). Исключая монозиготных близнецов, этот результат означает, что вероятность того, что любые два случайных NA-яка, случайно имеющих одинаковый генотип на всех 28 участках, составит 1 к 172 миллиардам. Этого достаточно для отслеживания яков и их продуктов в глобальной пищевой цепи, если это необходимо.
Вероятность исключения случайных взрослых из отцовства с 87 tySNP была высокой.У большинства случайных взрослых особей было от 5 до 10 сайтов tySNP, которые исключали их отцовство с данным потомством, то есть не имели общих аллелей с потомками на этих сайтах. Однако ложные исключения у истинного родителя из-за ошибки генотипа составляли только около 1 из 87 tySNP с частотой ошибок платформы генотипа 1%. Более 99% случайных взрослых могут быть исключены из отцовства даже при уровне ошибок генотипирования до 5% (т.е. четыре исключения разрешены для истинного родителя). Таким образом, большинство телят яков можно отнести к одному родителю без наличия генотипов другого родителя.Для ситуаций спаривания на пастбище с несколькими отцами это может снизить стоимость проверки отцовства почти на 50%. Обратите внимание, что эти оценки относятся к исключению отцовства среди взрослых, не являющихся родственниками. Возможность исключать полных сибсов из отцовства существенно снижена наполовину и, таким образом, может потребоваться использование генотипов матери и / или большей точности в системе генотипирования 9 . Когда задействовано 1000 потомков, одна из эффективных стратегий состоит в том, чтобы сначала установить высокоуверенные отношения между детенышами и потомками с помощью тестирования с одним родителем, а затем использовать генотипы матери для подтверждения исключения производителей из любых оставшихся неоднозначных отношений между детенышами и потомками.
Интрогрессия видов B. taurus у 170 яков NA из различных источников была низкой и не отличалась от таковой у других яков. Для 87 tySNP не было обнаружено значительных различий между средней частотой B. taurus -ассоциированного аллеля у 170 яков NA (0,0042) и девяти домашних и диких китайских яков (0,0057). Это предполагает, что генетическая основа популяции NA-яков в целом не подвергается значительному влиянию с интрогрессированной зародышевой плазмой B. taurus .Низкий фон интрогрессии крупного рогатого скота у яков способствует способности tySNP обнаруживать события гибридизации F1 у недавних предков яков. С 87 tySNP моделирование предсказало достоверное обнаружение событий гибридизации F1 у яков, содержащих всего лишь 1/16 часть зародышевой плазмы B. taurus (т.е. три поколения обратного скрещивания после события F1). Этого было бы достаточно для проверки точности родословных трех или четырех поколений. Однако следует отметить, что tySNP не информативны в отношении того, какой B.taurus участвовало в событии гибридизации F1, потому что почти все B. taurus животных являются гомозиготными по аллелю, ассоциированному с B. taurus . Мы также отметили, что шесть tySNP имели частоты аллелей, ассоциированных с B. taurus , между 0,01 и 0,10 у NA yak, в то время как частоты аллелей, ассоциированных с B. taurus , в другом 81 tySNP были практически нулевыми. Причины этого могут включать: интрогрессию и отбор древних B. taurus и / или наличие обоих аллелей до видообразования.Будущий фильтр tySNP, который следует учитывать, будет включать менее 1% распространенности B. taurus -ассоциированных аллелей у NA яков. Несмотря на эти исключения, в целом B. taurus -ассоциированные аллели в tySNP были редки у яков NA и, таким образом, были способны идентифицировать животных, происходящих от бэккроссов недавних гибридов F1.
Основные области для улучшения мультиплексного набора из 87 tySNP включают: изобилие маркеров, MAF по происхождению и геномное распределение. Представленные здесь 87 tySNP были разработаны из-за необходимости, до доступности сборки генома NA yak, и без предварительного знания частот их аллелей в популяциях NA.В то время как только 28 tySNP обеспечивают более чем достаточную мощность для генетического дактилоскопирования, дополнительные tySNP могут добавить значительную силу для исключения отцовства. Использование будущих WGS от дополнительных 10–15 яков NA позволит уверенно присвоить оценки MAF для 610 tySNP, представленных здесь. Имея эту информацию о частоте аллеля NA yak, выбор tySNP с наивысшими MAF и увеличение количества маркеров по крайней мере до 131 (т. Е. 1,5x) увеличит долю случайных взрослых, исключенных из отцовства, как показано на рисунке 7B.Это также помогло бы исключить высокородственных животных из отцовства, например, когда полные однояйцевые быки одновременно контактируют с коровами. Наличие набора из 131 tySNP с высоким MAF также несколько улучшит способность обнаруживать события гибридизации F1. Моделирование показало, что для получения еще одного поколения чувствительности для обнаружения гибридизации F1 потребуется 175 (т.е. 2x) tySNP. По мере совершенствования технологий генотипирования и повышения их рентабельности может появиться возможность включать дополнительные маркеры из 421 бункера 610 tySNP.
Триаллельные SNP были систематически идентифицированы у других млекопитающих. В Homo sapiens набор из 1270 полиморфных триаллельных SNP, добытых в рамках проекта 1000 Genomes Project, недавно был использован для судебно-медицинской идентификации пропавших без вести 34 . Интересно, что у людей существует примерно в два раза больше триаллельных сайтов, чем ожидалось случайно 35 . Вполне возможно, что по крайней мере некоторые из них могут быть связаны с интрогрессией архаичных видов в современных людей 36 .Независимо от их использования на людях, представленный здесь многоцелевой триаллельный подход SNP может быть полезен для других видов, которые могут гибридизоваться, включая крупного рогатого скота, цервид, зубатых, верблюжьих, непарнокопытных, псовых, кошачьих и медвежьих. Очевидное непосредственное применение — у равнинных бизонов Северной Америки ( B. bison ). Равнинные бизоны — дикие животные, обитающие в Северной Америке, столкнулись с проблемой ограничения популяции, скрещиваются с B. taurus и выращиваются для получения мяса и побочных продуктов. В идеале, WGS из разнообразной группы из 15 неродственных равнинных бизонов, отобранных из стада NA, должны использоваться вместе с бантенгами, гаурами, яками и рогатым скотом для идентификации подходящих триаллельных SNP бизонов (tbSNP).При использовании WGS от 15 бизонов оценки MAF для происхождения бизонов были бы известны как априори и могли бы более эффективно использоваться в процессе фильтрации tbSNP. В зависимости от целей, WGS древесного бизона NA ( B , bison athabascae ) или европейского зубра ( B. bonasus ) также могут быть включены в фильтрацию для оценок MAF и расширить потенциальную полезность панели tbSNP.
Заключение
Представленные здесь результаты новых tySNP демонстрируют, что один минимальный набор маркеров может эффективно и точно использоваться для идентификации животных, определения происхождения и обнаружения недавних событий гибридизации F1 для поддержки управления стадом и решений производителей яков о разведении.610 tySNP, информация о дизайне их анализа, множественные тесты MALDI-TOF MS для подмножества 87 tySNP и вся другая связанная информация доступна для использования во всем мире без ограничений.
Доступность данных
Базовые данные
NCBI BioProject: Bos mutus штамм: yakQh2 Секвенирование и сборка генома. Регистрационный номер PRJNA74739.
NCBI BioProject: Bos grunniens Секвенирование генома. Регистрационный номер PRJNA285834.
NCBI BioProject: Поэтапная трио сборка геномов яка и крупного рогатого скота.Регистрационный номер PRJNA551500.
NCBI BioProject: Bos grunniens x Bos taurus Секвенирование и сборка генома. Регистрационный номер PRJNA552915.
NCBI BioProject: данные WGS по различным типам крупного рогатого скота США. Регистрационный номер PRJNA324822.
NCBI BioProject: данные WGS от Cetartiodactyla. Регистрационный номер PRJNA325061.
Доступность программного обеспечения
Все специализированное программное обеспечение, используемое для анализа данных для этого проекта, доступно на странице GitHub этого проекта: https: // github.com / kalbflei / YakParentageAndIntrogression.
Программное обеспечение, заархивированное на момент публикации: https://doi.org/10.5281/zenodo.3988457 28 .
Лицензия: Лицензия MIT.
snps · PyPI
snps стремится быть простой в использовании и доступной библиотекой с открытым исходным кодом для работы с данные генотипа
Примеры
Загрузить данные примера
Во-первых, давайте настроим ведение журнала, чтобы получить полезный результат:
>>> импорт журнала, sys >>> logger = ведение журнала.getLogger () >>> logger.setLevel (logging.INFO) >>> logger.addHandler (logging.StreamHandler (sys.stdout))
Теперь мы готовы загрузить несколько примеров данных из openSNP:
>>> from snps.resources import Resources >>> r = Ресурсы () >>> пути = r.download_example_datasets () Качаем ресурсы / 662.23andme.340.txt.gz Ресурсы для скачивания / 662.ftdna-illumina.341.csv.gz
Загрузить необработанные данные
Загрузить файл исходных данных 23andMe:
>>> from snps import SNPs
>>> s = SNP ("ресурсы / 662.23andme.340.txt.gz ")
>>> s.source
'23andMe'
>>> s.count
9
Класс SNP принимает путь к файлу или байтовому объекту. Класс Reader пытается вывести источник данных и загрузить SNP. Загруженные SNP нормализованные и доступно через pandas.DataFrame:
>>> df = s.snps >>> df.columns.values array (['chrom', 'pos', 'genotype'], dtype = объект) >>> df.index.name 'rsid' >>> df.chrom.dtype.name 'объект' >>> df.pos.dtype.name 'uint32' >>> df.genotype.dtype.name 'объект' >>> len (df) 9
snps также пытается обнаружить сборку / сборку данных:
>>> s.build 37 >>> s.build_detected Истинный >>> s.assembly 'ГРЧ47'
Объединить файлы сырых данных
Набор данных состоит из файлов необработанных данных из двух разных источников тестирования ДНК — давайте объединим эти файлы. В частности, мы обновим объект SNP с помощью SNP из Файл ДНК семейного древа.
>>> merge_results = s.merge ([SNP ("resources / 662.ftdna-illumina.341.csv.gz")])
Объединение SNP ('resources / 662.ftdna-illumina.341.csv.gz')
SNP ('resources / 662.ftdna-illumina.341.csv.gz') имеет сборку 36; переназначение на сборку 37
Ресурсы для скачивания / NCBI36_GRCh47.tar.gz
27 позиций SNP не совпадали; сохранение исходных позиций
151 генотип SNP не совпал; пометить их как нулевые
>>> s.source
'23andMe, FTDNA'
>>> s.count
1006960
>>> s.build
37
>>> с.build_detected
Истинный
Если объединяемые SNP имеют сборку, которая отличается от целевой сборки, SNP для объединения будет автоматически переназначен. После слияния этого примера сборка все еще обнаруживается, поскольку build обнаружен для всех объединенных объектов SNP.
По мере добавления данных они сравниваются с существующими, а также с положением и генотипом SNP. Выявлены несоответствия. (Пороги несоответствия можно настроить с помощью параметров.) несоответствующие SNP доступны для проверки после слияния через свойства объекта SNP.
>>> len (s.discrepant_merge_genotypes) 151
Кроме того, любые неназванные / нулевые генотипы будут обновлены во время слияния, если файл объединяемый имеет так называемый генотип для SNP.
Кроме того, слияние принимает параметр chrom — это позволяет объединять только связанные SNP. с указанной хромосомой (например, «Y» или «MT»).
Наконец, merge возвращает список dict, где каждый dict имеет информацию, соответствующую к результатам каждого слияния (например,g., общие SNP).
>>> отсортировано (список (merge_results [0] .keys ())) ['common_rsids', 'Dispant_genotype_rsids', 'Dispant_position_rsids', 'объединенный'] >>> merge_results [0] ["объединено"] Истинный >>> len (merge_results [0] ["common_rsids"]) 692918
Переназначить SNP
Теперь давайте повторно сопоставим объединенные SNP, чтобы изменить сборку / сборку:
>>> s.snps.loc ["rs3094315"]. Pos 752566 >>> chromosomes_remapped, chromosomes_not_remapped = s.переназначить (38) Ресурсы для скачивания / GRCh47_GRCh48.tar.gz >>> s.build 38 >>> s.assembly 'ГРЧ48' >>> s.snps.loc ["rs3094315"]. pos 817186
SNP можно переназначить между сборкой 36 (NCBI36), сборкой 37 (GRCh47) и сборкой 38 (ГРЧ48).
Сохранить SNP
Хорошо, пока мы объединили SNP из двух файлов (обеспечивая одинаковую сборку в процессе и выявление несоответствий по ходу дела). Затем мы переназначили SNP на сборку 38. Теперь давайте сохраним объединенный и переназначенный набор данных, состоящий из 1M + SNP, в файл значений, разделенных табуляцией (TSV):
>>> сохранено_snps = s.сохранить ("out.txt")
Сохранение output / out.txt
>>> печать (сохраненные_snps)
вывод / out.txt
Кроме того, давайте получим ссылочные последовательности для этой сборки и сохраним SNP в виде файла VCF:
>>> saved_snps = s.save ("out.vcf", vcf = True)
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.1.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.2.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.3.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.ГРЧ48.dna.chromosome.4.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.5.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.6.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.7.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.8.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.9.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.10.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.ГРЧ48.dna.chromosome.11.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.12.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.13.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.14.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.15.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.16.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.17.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.18.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.19.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.20.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.21.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.22.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.X.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.Y.fa.gz
Ресурсы для скачивания / fasta / GRCh48 / Homo_sapiens.GRCh48.dna.chromosome.MT.fa.gz
Сохранение output / out.vcf
1 Обнаружено несоответствие позиций SNP при сохранении VCF
При сохранении VCF, если какие-либо SNP имеют позиции вне ссылочной последовательности, они помечаются как несоответствующие и доступны через свойство объекта SNP.
Все выходные файлы сохраняются в выходной каталог.
»Аккредитованные лаборатории для проверки отцовства крупного рогатого скота по SNP и STR
Аккредитованные лаборатории для проверки отцовства крупного рогатого скота с помощью SNP и STR
На этой странице представлены актуальные аккредитованные лаборатории для проверки отцовства крупного рогатого скота с помощью SNP и STR.
Страница с инструкциями по предоставлению аккредитации ICAR доступна здесь
Секретариат напоминает, что аккредитация ICAR действительна в течение двух лет
лабораторий аккредитованы в 2020 г. (декабрь). Аккредитация по «тестированию отцовства на основе SNP» и «тестированию отцовства на основе STR» у крупного рогатого скота действительна до декабря 2022 года, если не указано иное
| Тип аккредитации (срок действия до декабря 2022 года) | ||||
| Название лаборатории | Главный офис | Контактное лицо и электронная почта | «Тестирование отцовства на основе SNP» крупного рогатого скота | «Тестирование отцовства на основе STR» крупного рогатого скота |
| Аверон Лаб | Франция | Роланд Бругиду | Аккредитованный | Аккредитованный |
| Служба селекции Словацкой Республики — Лаборатория генетики | Словацкая респ. | Моника Фецкова | Аккредитованный | |
| TERANA Nièvre (ранее: Laboratoire Départmental d’Analyses et de Conseil de la Nièvre) | Франция | Шанталь Одеваль | Аккредитованный | Аккредитованный |
| Чешско-моравская селекционная корпорация, Лаборатория иммуногенетики | Чешская Республика | Михаэла Прибанова | Аккредитованный | Аккредитованный |
| Лабогена ДНК | Франция | Ив Агез | Аккредитованный | Аккредитованный |
| Национальный научно-исследовательский институт животноводства — лаборатория геномики | Польша | Катаржина Ропка-Молик | Аккредитованный | |
| LGS — Agrotis | Италия | Раффаэле Мазза | Аккредитованный | |
| Неоген | США | Цзяньшэн Цю | Аккредитованный | Аккредитованный |
| ULG GIGA-Genomics | Бельгия | Воутер Коппитерс | Аккредитованный | |
| Weatherbys Scientific | Ирландия | Эйдан Мартин | Аккредитовано до января 2021 г. с продлением аккредитации, предоставленной в 2018 г. | |
| Xenética Fontao S.А. | Испания | Хавьер Лопес Виана | Аккредитованный | |
Лаборатории, аккредитованные в 2019 г. (декабрь), чья аккредитация действительна до декабря 2021 г., путем «проверки отцовства на основе SNP» и «проверки отцовства на основе STR» для крупного рогатого скота
| Тип аккредитации (срок действия до декабря 2021 года) | ||||
| Название лаборатории | Главный офис | Контактное лицо и электронная почта | «Тестирование отцовства на основе SNP» крупного рогатого скота | «Тестирование отцовства на основе STR» крупного рогатого скота |
| Агранис | Франция | Флоран Перрен | Аккредитованный | |
| Certagen | Германия | Юрген Вебер | Аккредитованный | |
| Чешско-моравская селекционная корпорация, Лаборатория иммуногенетики | Чешская Республика | Михаэла Прибанова | Аккредитованный | |
| Dr. van Haeringen Laboratorium b.v. | Нидерланды | Линн ван де Гур | Аккредитованный | |
| GeneControl | Германия | Дорис Зайхтер | Аккредитованный | Аккредитованный |
| Диффузия генов | Франция | Пьер Бувель | Аккредитованный | |
| Лаборатория молекулярной генетики | Польша | Катаржина Ропка-Молик | Аккредитованный | |
| Институт зоотехники Маэбаши | Япония | Ёсихиро Танабэ | Аккредитованный | Аккредитованный |
| Неоген | США | Цзяньшэн Цю | Аккредитованный | Аккредитованный |
| Проген | Бельгия | Томас Валери | Аккредитованный | |
| Tierärztliches Institut Uni Göttingen | Германия | Бертрам Брениг | Аккредитованный | Аккредитованный |
| ULG GIGA-Genomics | Бельгия | Воутер Коппитерс | Аккредитованный | |
| Zoetis Genetics | США (Каламазу, Мичиган) | Карен Резерфорд | Аккредитованный | Аккредитованный |