Расшифровка фбс блоки: что это такое? Расшифровка ФБС
Блок бетонный для стен подвалов ФБС24.5.6-Т
Расшифровка аббревиатуры ФБС – фундаментные блоки сплошные. Это говорит о том, что в отличие от пустотелых аналогов, данные изделия характеризуются монолитной структурой, что позволяет им выдерживать значительные нагрузки.
Область применения
Как говорилось выше, блоки фбс 24 5 6 применяются в нагруженных конструкциях. Чаще всего их используют для обустройство ленточных фундаментов. Множество жилых многоэтажных домов в городе Москва было построено с применением блоков данного типа. При этом, такие изделия позволяют строить дома как с подвальными помещениями, так и без них. А потому, еще одна сфера их применения – это строительство стен и перегородок в подвальных помещениях.
В отдельных случаях блок фбс 24 5 6 может использоваться для строительства небольших хозпостроек и гаражных помещений.
Технология производства и сырье
Основным критерием в изготовлении блоков ФБС является качество бетона.
В процессе изготовления блоков используется целый ряд спецоборудования, которое обеспечивает высокую прочность конечного продукта:
- Обычные формы и виброформы.
- Глубинные вибраторы.
- Бетономешалки.
Если же говорить о процессе производства ФБС блоков, то он начинается с изготовления смеси из цемента, воды и специального наполнителя. При этом, четко соблюдаются необходимые дозировки. Далее, когда состав перемешается в бетономешалке, его разливают по формам для прохождения вибропрессования. Уплотнение бетона осуществляется с применением глубинного вибратора.
Через сутки плиты извлекаются из форм, после чего они должны набирать прочность в течение недели. Окончательной прочности бетон достигает по истечении 28 дней.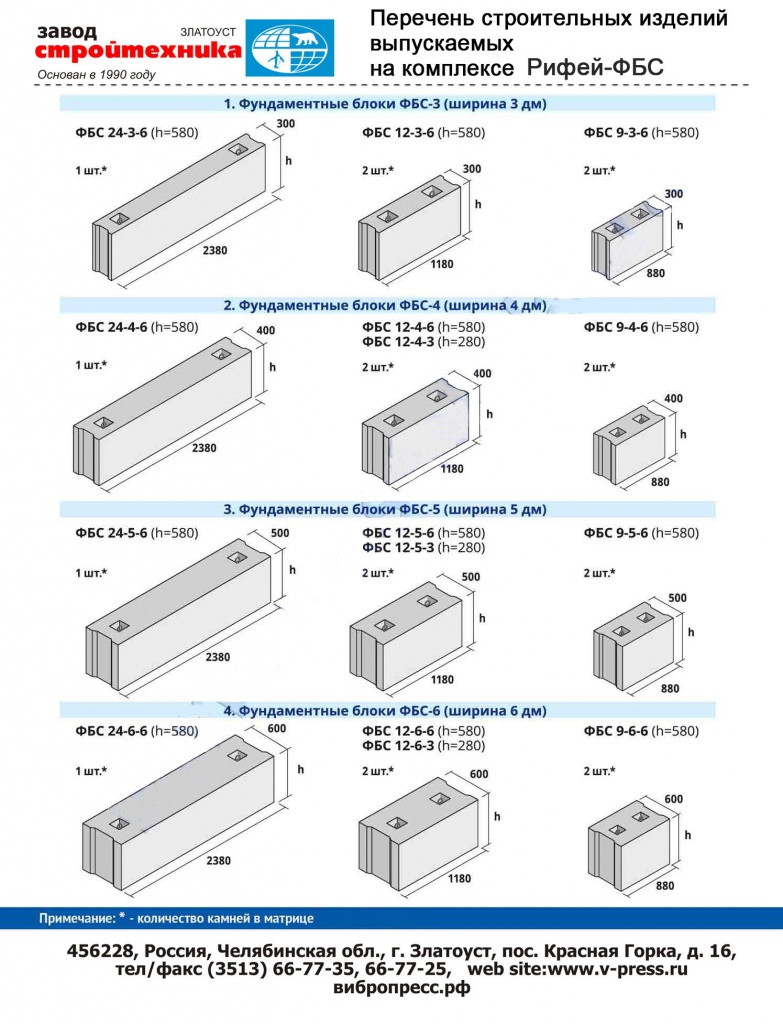
Как хранить и транспортировать
Хранить блок фбс 24 5 6 т необходимо в вертикальном положении, обязательно на складе. Каждый блок устанавливается на специальную инвентарную подкладку, толщиной не менее 3 см. Между штабелями должно сохраняться достаточное расстояние для того, чтобы перемещать их посредством подъемного механизма. Штабеля не должны превышать
Для транспортировки блоки прочно укрепляются в кузове транспортного средства. При этом, крепления должны исключать возможность смещений в процессе транспортировки. Погрузка-выгрузка осуществляется только спецтехникой.
Как заказать
Если вы хотите заказать блоки ФБС в Москве, то компания ЖБИКОМ – это именно то, что вам нужно. Здесь вы сможете приобрести качественные блоки фбс 24 5 6 цена которых является более, чем приемлемой. Вашему вниманию также предлагается доставка по Москве и возможность осуществить заказ любым удобным для вас способом:
- По телефону.

- Посредством заполнения формы на сайте.
- Отправив электронное письмо.

Доверяйте профессионалам и приобретайте качественные стройматериалы!
Фундаментные блоки, или ФБС
Фундаментные блоки, или ФБС – это один из наиболее востребованных строительных материалов, позволяющий быстро соорудить надежный и долговечный фундамент. Производят ФБС из высокопрочных тяжёлых бетонов, укрепленных стальной арматурой, поэтому они отличаются долговечностью и способны длительное время выдерживать интенсивные высокие нагрузки. Благодаря высокому качеству бетона блоки в процессе эксплуатации не подвергаются деформации, легко выдерживают суровые морозы, при грамотной гидроизоляции не боятся влаги.
Традиционная форма ФБС – параллелепипед, но сегодня производители предлагают блоки самых разных конфигураций. Это позволяет воплощать в реальности самые оригинальные и смелые архитектурные проекты. Кроме монтажа фундаментов, ФБС блоки активно используют при возведении подвальных помещений, ограждений, а также стен промышленных зданий и других объектов, подверженных высоким нагрузкам и требующих повышенной прочности.
Кроме монтажа фундаментов, ФБС блоки активно используют при возведении подвальных помещений, ограждений, а также стен промышленных зданий и других объектов, подверженных высоким нагрузкам и требующих повышенной прочности.
Преимущества применения ФБС
Использование фундаментных блоков при возведении фундаментов дает следующие преимущества: возможность проведения монтажных работ всесезонно и при любой погоде; монтаж блоков гораздо проще, чем заливка фундамента, это позволяет значительно сократить сроки строительства; их использование для сооружения фундамента позволяет равномерно распределить нагрузки по всей площади основания; повышенная надежность фундамента.
Перед тем, как выбрать ту или иную разновидность фундаментных блоков, на участке, где планируется строительство, проводят инженерно-геологические изыскания для определения типа грунта и расчета ожидаемых нагрузок на фундамент.
Виды ФБС
Фундаментные блоки бывают трех типов:
- сплошные;
- с вырезами для прокладки инженерных коммуникаций и монтажа перемычек;
- пустотные.

Что собой представляют блоки ФБС, их назначение
Любая аббревиатура скрывает за сухим обозначением определённое понятие. ФБС, если обратиться к дословной расшифровке — это фундаментные блоки строительные. Причём по умолчанию подразумевается, что такая маркировка означает именно железобетонную конструкцию прямоугольной формы без каких-либо полостей внутри, то есть цельную. Говоря более простым языком, такие блоки — не что иное, как привычный многим строительный материал кирпич, но усиленный арматурным каркасом, так называемые «подушки», которые обозначаются как ФЛ.
ФБС блоки состоят из нижней и верхней панелей, соединенных продольной стенкой и стойками. Поперечное сечение блоков может быть трапецеидальным либо тавровым. Процесс производства ФБС регламентируется государственными стандартами. В маркировке блоков содержится информация о параметрах ЖБ изделий — высота, ширина и длина. Значения в маркировке приводятся в дециметрах.
При выборе фундаментных блоков необходимо учитывать такие факторы, как:
- этажность возводимого объекта;
- тип грунта, на котором сооружается фундамент;
- параметры блоков;
- их технические характеристики, включая влагопроницаемость и морозостойкость.

Монтаж фундаментных блоков
Для монтажа готовых бетонных блоков как правило применяется автокран либо аналогичные подъемные механизмы. Исключения составляют лишь мелкоразмерные железобетонные блочки весом до 100 кг, которые можно монтировать вручную. Рассмотрим два основных варианта монтажа фундаментов из сборных ЖБ элементов.
Вариант с независимым полом подвала, заливаемым после монтажа основного фундамента. Сначала разрабатывается котлован. В основании котлована делается песчаная подушка. По всей площади котлована, если планируется обустраивать подвальное помещение. Либо непосредственно по периметру внешних и внутренних стен, если не будет отливаться бетонный пол подвала. На дне котлована, на уложенные фундаментные подушки ФЛ (фундаменты ленточные) монтируются фундаментные бетонные блоки ФБС.
Монтаж бетонных блоков ведётся в разбежку, подобно кирпичной кладке, на цементный раствор, в четыре-пять рядов в высоту.
Грамотную гидроизоляцию тоже не сделать, всё из-за той же независимой подвижки пола и стен. Возможны варианты с анкеровокй. Т.е. продольная и поперечная арматура будущей плиты пола подвала загоняется своими концами в стены. Для этого, лента из блоков засверливается перфоратором по всему периметру будущей плиты. В эти отверстия и заводят хвосты арматуры. В таком случае плита должна быть не тоньше 15-20 см. Однако, делать это в первый год жизни здания — всё же не стоит. Усадка ленты фундамента может нарушить целостность плиты, либо просто срежет заанкерённые хвостовики арматуры. Если речь идёт об анкеровке стандартной (в бытовом строительстве) арматурой 10-12 мм. С арматурой 16-20 мм такого казуса, скорее всего, не произойдёт. В подвалах зданий, прошедших основную стадию усадки первого года, полы можно отливать без анкеровки. Но всё же более предпочтителен другой вариант.
Т.е. продольная и поперечная арматура будущей плиты пола подвала загоняется своими концами в стены. Для этого, лента из блоков засверливается перфоратором по всему периметру будущей плиты. В эти отверстия и заводят хвосты арматуры. В таком случае плита должна быть не тоньше 15-20 см. Однако, делать это в первый год жизни здания — всё же не стоит. Усадка ленты фундамента может нарушить целостность плиты, либо просто срежет заанкерённые хвостовики арматуры. Если речь идёт об анкеровке стандартной (в бытовом строительстве) арматурой 10-12 мм. С арматурой 16-20 мм такого казуса, скорее всего, не произойдёт. В подвалах зданий, прошедших основную стадию усадки первого года, полы можно отливать без анкеровки. Но всё же более предпочтителен другой вариант.
Вариант с монолитной плитой в основании. Разрабатывается котлован. По всей площади дна котлована делают песчаную подушку. Во избежание непредсказуемых усадок, уложенный песок проливают водой, затем его тщательно утрамбовывают виброплитой. В идеальном варианте стоит сразу позаботиться о гидроизоляции. Для этого, на утрамбованную песчаную подушку отливается предварительная плита из низкомарочного бетона м-100, толщиной примерно 7-10 см. Это так называемая подготовка. На неё укладывают рулонную гидроизоляцию (гидростеклоизол) и тщательно спаивают. Оставляя выпуски во все стороны по 50-100 см. Далее по всей площади котлована изготовляется арматурный каркас (поверх гидроизола). Ставится мини-опалубка по внешнему периметру будущей плиты. Плита может быть несколько больше в ширину и длину, чем будущий фундамент. Как бы с напуском в стороны. Впрочем, это не обязательно.
В идеальном варианте стоит сразу позаботиться о гидроизоляции. Для этого, на утрамбованную песчаную подушку отливается предварительная плита из низкомарочного бетона м-100, толщиной примерно 7-10 см. Это так называемая подготовка. На неё укладывают рулонную гидроизоляцию (гидростеклоизол) и тщательно спаивают. Оставляя выпуски во все стороны по 50-100 см. Далее по всей площади котлована изготовляется арматурный каркас (поверх гидроизола). Ставится мини-опалубка по внешнему периметру будущей плиты. Плита может быть несколько больше в ширину и длину, чем будущий фундамент. Как бы с напуском в стороны. Впрочем, это не обязательно.
Отливается плита из бетона от М-200 до М-300. Далее, на эту плиту монтируются внешние и внутренние стены подвала из фундаментных блоков ФБС. Всё так же как и в первом варианте. Стены так же обклеивают рулонной гидроизоляцией и склеивают её горелкой или паяльной лампой с нижним слоем гидростеклоизола, которую уложили на подготовку.
Основные преимущества данного варианта:
- Целостность основания
- Равномерная усадка всего фундамента и здания. Без возможных проседаний и перекосов.
- Жёсткая конструкция стены — пол, позволяющая сделать качественную гидроизоляцию и отделку.
- Увеличенная площадь опоры, и как следствие — лучшая несущая способность фундамента.
 Без возможных проседаний и перекосов.
Без возможных проседаний и перекосов.
Перечисленных преимуществ — более чем достаточно, чтобы сделать выбор в пользу варианта с нижней плитой. Безусловно, не стоит забывать про гидроизоляцию стен и пола фундамента, при этом используют бетоны и растворы на специальных видах цемента. Для внешней защиты стен из фундаментных блоков, часто применяют обмазочную или обклеечную гидроизоляцию материалами на основе битума. Внутреннюю гидроизоляцию — различными гидрофобизаторами и пропитками типа Кальматрона, двухкомпонентными обклеечными составами и т.д. В ситуации с высоким уровнем грунтовых вод, и опасностью сезонного подтопления подвалы из сборного железобетона уступают своим монолитным собратьям. Блоки фундаментные ФБС помимо своего основного предназначения, могут так же применяться при сооружении технических зданий, гаражей, складов и так далее.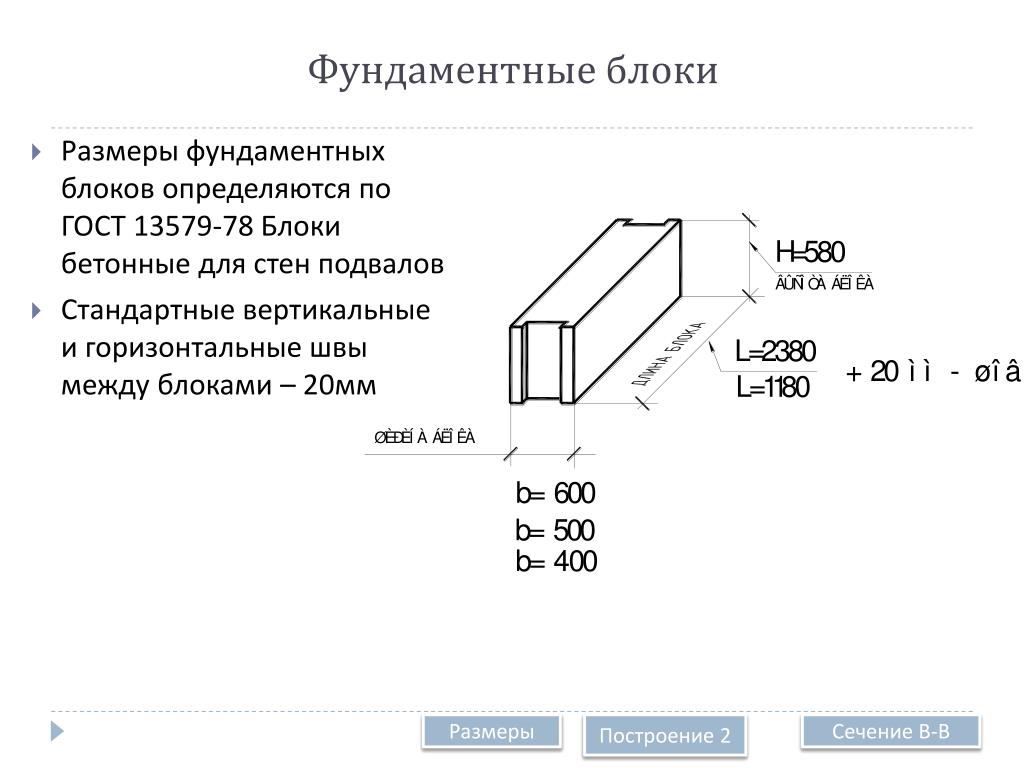 Часто ФБС-ки используют в качестве временных ограждений от въезда автомобилей и т.д. Безусловно они являются одним из самых распространённых изделий ЖБИ.
Часто ФБС-ки используют в качестве временных ограждений от въезда автомобилей и т.д. Безусловно они являются одним из самых распространённых изделий ЖБИ.
Каковы достоинства и недостатки блоков ФБС
Когда планируется возведение постройки на основе такой конструкции, как фундамент из блоков ФБС, все имеющиеся плюсы и минусы необходимо тщательно проработать перед тем, как склониться в сторону этого или другого варианта. Привлекателен данный вид основы под постройку по целому ряду причин:
- Экономия времени, так как отсутствует необходимость замешивать бетон, рассчитывать количество необходимого материала, а также заботиться о соблюдении идеальных пропорций. Такое случается часто, если нет опыта в этой области деятельности. А укладка блоков осуществляется быстро и не придётся опасаться по поводу качества будущего фундамента.
- Производителем предусмотрены практически все нюансы при возведении основы под постройку из блоков ФБС. Речь идёт о специальных крюках, которыми оснащены изделия особо крупных габаритов. При этом намного упрощается процесс укладки таких изделий в ямы при строительстве многоэтажных или просто тяжёлых конструкций.
- Различные размеры блоков позволяют построить необходимый по размеру фундамент или другой объект.
- Если строительство планируется на «кислых» грунтах или в местности с очень морозным климатом, то благодаря своей структуре блоки ФБС не поддаются воздействию перечисленных внешних факторов.
 При этом намного упрощается процесс укладки таких изделий в ямы при строительстве многоэтажных или просто тяжёлых конструкций.
При этом намного упрощается процесс укладки таких изделий в ямы при строительстве многоэтажных или просто тяжёлых конструкций.Разбираясь, какие же недостатки имеются у данных блоков можно выделить два существенных. Во-первых, для укладки придется использовать специальную технику. А во-вторых, есть риск проседания фундамента из-за того, что конструкция не монолитная, а значит, каждый из элементов может повести себя непредсказуемо.
Некоторые приписывают к недостаткам ФБС необходимость дополнительной гидроизоляции и утепления, но ведь сегодня практически каждый из видов оснований под постройку не обходится без таких манипуляций. Соответственно, считать это минусом именно в случае блоков ФБС ошибочно.
Соответственно, считать это минусом именно в случае блоков ФБС ошибочно.
Таким образом, выбирая фундамент из железобетонных фундаментных блоков при возведении постройки, можно не опасаться ни перепадов температур, ни прочих воздействий внешних факторов. А степень надёжности зданий, построенных именно на таком фундаменте, крайне высока. Плюс ко всему — конструкция из блоков возводится очень быстро, а значит, нет повода переживать о сроках строительства и можно успеть закончить работы в промежуток между сменами сезонов года. Даже если планируется построить одноэтажный жилой дом скромных габаритов, выбрав такой вид фундамента можно получить строение «на века».
ФБС 9.4.6-Т
Железобетонные изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов – это несущие опорные конструкции, которые широко используются в строительстве зданий и сооружений. Высокопрочные сборные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов выполняют функцию основания, на котором возводятся малоэтажные здания и промышленные строения различного назначения.
Железобетонные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов широко используются при возведении столбчатых и ленточных фундаментов. Бетонные блоки укладываются по периметру и осевым линиям проектируемого сооружения, а стык между ними заливается специальным раствором, обеспечивающим прочность соединения. Фундамент из сборных бетонных блоков воспринимает нагрузки, которые передаются от веса и стен здания, и распределяет их далее в грунт.
Кроме того, с помощью фундаментных блоков строятся стены подвальных или цокольных помещений различной глубины, а также технические подполья зданий. Основное направление, где применяются фундаменты из сборных блоков – это промышленное строительство. Однако небольшой вес блоков позволяет использовать их в частном строительстве: без необходимости привлечения тяжелой техники можно в кратчайшие сроки устроить ленточный фундамент любой площади и конфигурации. Изделия фундаментные блоки сплошные ФБС 9.4. 6-Т из тяжелого бетона для стен и подвалов, будучи универсальными ЖБИ, используются и в дорожном строительстве, где они выступают в роли барьерных преград и невысоких ограждений на проезжей части автомобильных дорог, а также в местах, не предназначающихся для проезда автотранспорта.
6-Т из тяжелого бетона для стен и подвалов, будучи универсальными ЖБИ, используются и в дорожном строительстве, где они выступают в роли барьерных преград и невысоких ограждений на проезжей части автомобильных дорог, а также в местах, не предназначающихся для проезда автотранспорта.
Сборные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов из бетона обладают достаточной прочностью для укладки в любых типах грунтов и в любых климатических условиях. Необходимый уровень влагостойкости и морозостойкости достигается добавлением специальных химических присадок, которые добавляются в бетонную смесь при производстве изделий и стык между блоками при их установке на стройплощадке. Изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов обладают важнейшим преимуществом по сравнению с монолитными фундаментами: стоимость отдельных блоков и их монтаж обходятся значительно дешевле, а простота монтажа и возможность его проведения в любое время года позволяет сэкономить время, затрачиваемое на строительство сооружений.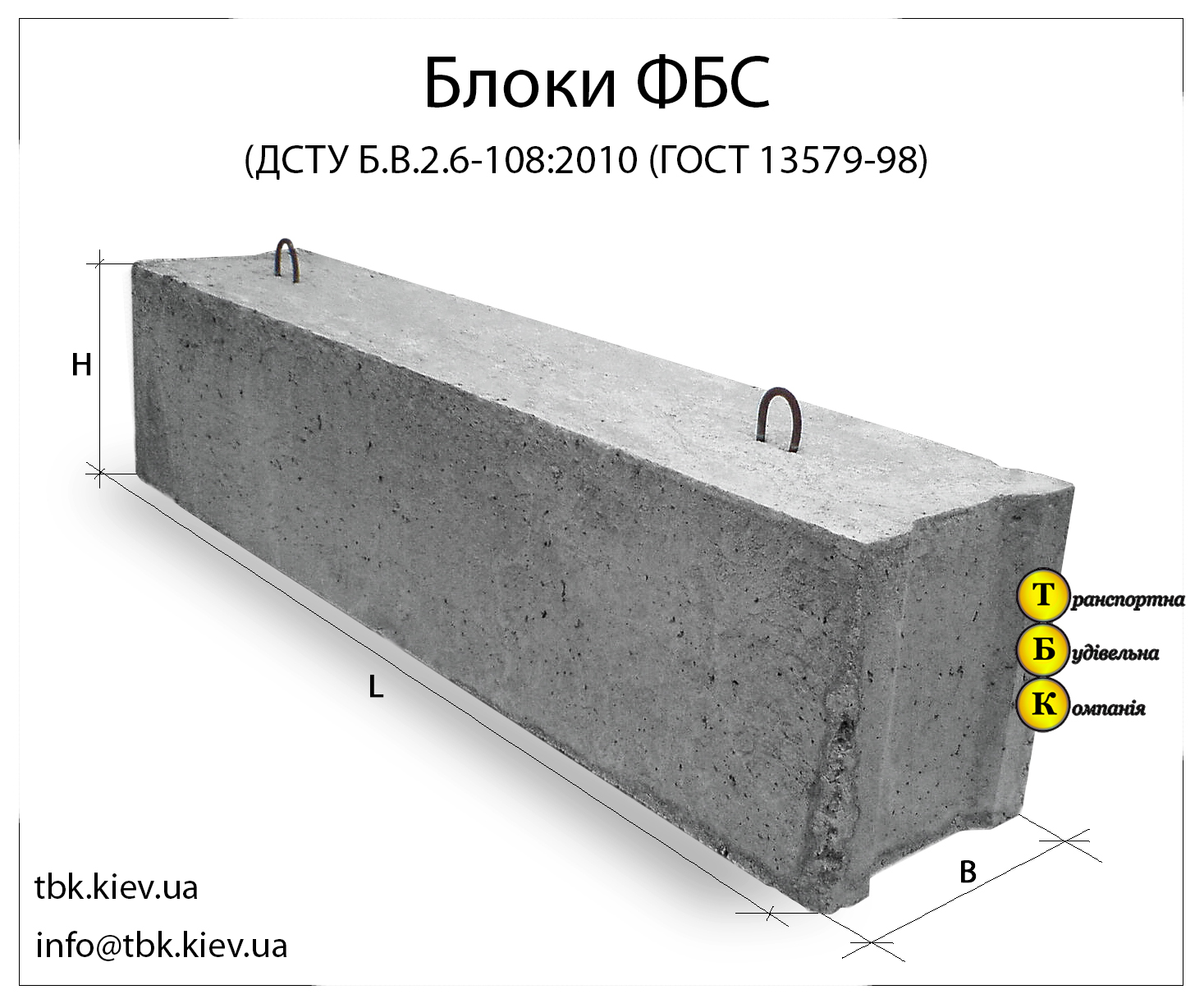 Помимо этого, в случае поломки изделия нет необходимости целиком ремонтировать фундамент, достаточно лишь заменить поврежденный элемент.
Помимо этого, в случае поломки изделия нет необходимости целиком ремонтировать фундамент, достаточно лишь заменить поврежденный элемент.
Изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов представляет собой прямоугольный параллелепипед, боковые грани которого имеют специальные пазы, предназначенные для стыка блоков друг с другом. Пазы заполняются специальным цементным раствором, который обеспечивает необходимую прочность соединения и защиту от негативного влияния окружающей среды. На верхней плоскости фундаментные блоки имеют подъемные петли, служащие для удобства монтажа. При использовании специальных захватных устройств для подъема и монтажа блоков допускается изготовление блоков без подъемных петель.
Железобетонные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов изготавливаются в соответствии с нормами и указаниями, данными в ГОСТ 13579-78 «Блоки бетонные для стен подвалов» из тяжелого, легкого или силикатного бетона. Класс бетона по прочности на сжатие принимается не менее В3,5 и не более В15 и – соответственно, для блоков из легкого и тяжелого бетонов, и В12,5 – для блоков из плотного силикатного бетона. Классы по морозостойкости и водонепроницаемости бетона назначаются индивидуально для каждого проекта в зависимости от режима эксплуатации конструкций и климатических условий в районе строительства.
Класс бетона по прочности на сжатие принимается не менее В3,5 и не более В15 и – соответственно, для блоков из легкого и тяжелого бетонов, и В12,5 – для блоков из плотного силикатного бетона. Классы по морозостойкости и водонепроницаемости бетона назначаются индивидуально для каждого проекта в зависимости от режима эксплуатации конструкций и климатических условий в районе строительства.
Изделия ЖБИ фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов, как правило, не армируются, однако в некоторых случаях блоки могут иметь арматурные выпуски для усиления прочности фундамента: блоки жестко связываются между собой и объединяются в единую рамную конструкцию. Подъемные петли, с помощью которых фундаментные блоки транспортируются и монтируются, изготавливаются из стальной стержневой горячекатаной гладкой арматуры класса А-I или из стали периодического профиля Аc-II.
Железобетонные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов – это опорные конструкции, которые воспринимают значительные усилия.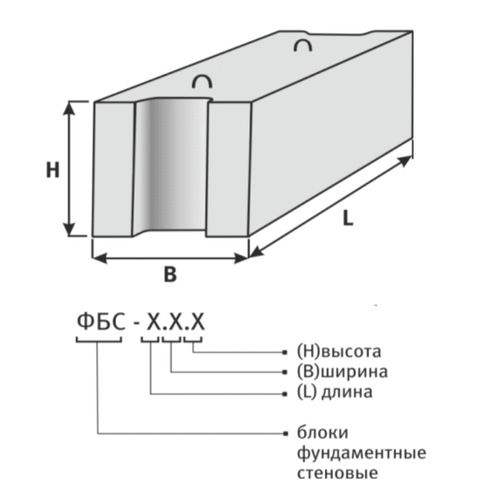 От качества их изготовления зависит надежность и долговечность построек. Поэтому к изделиям предъявляются достаточно серьезные требования: блоки фундаментные подвергаются контрольным испытаниям на каждом этапе производства с целью предотвращения брака, который может привести не только к проседанию и обрушению строения, но и к угрозе жизни человека.
От качества их изготовления зависит надежность и долговечность построек. Поэтому к изделиям предъявляются достаточно серьезные требования: блоки фундаментные подвергаются контрольным испытаниям на каждом этапе производства с целью предотвращения брака, который может привести не только к проседанию и обрушению строения, но и к угрозе жизни человека.
Данные изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов в момент отпуска с завода подлежат обязательной диагностике по прочности бетона. Значение фактической отпускной прочности бетона изделий принимается: 50% от класса по прочности на сжатие – для тяжелого и легкого бетонов класса В12,5 и выше; 70% – для тяжелого бетона класса В10 и ниже; 80% – для легкого бетона класса В10 и ниже; 100% – для плотного силикатного бетона. Поставка ЖБИ блоков с отпускной прочностью бетона ниже прочности, соответствующей его классу по прочности на сжатие, допускается производить только при условии, если изготовитель гарантирует достижение бетоном требуемой прочности в проектном возрасте.
Изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов не допускают трещин, кроме местных поверхностных усадочных, ширина которых не должна превышать 0,1-0,2 мм. Допускаются незначительные отклонения проектных размеров, значения которых не должны превышать 1±3 мм – по длине изделий, ±8 мм – по ширине и высоте, ±5 мм – по размерам пазовых вырезов. Отклонение от прямолинейности профиля поверхностей блока не должно превышать 3 мм по всей длине и ширине блока. Монтажные петли должны быть в обязательном порядке очищены от наплывов бетона.
Железобетонные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов хранятся в штабелях, рассортированные по маркам и партиям и уложенные вплотную друг к другу. Высота штабеля принимается не более 2,5 м. При хранении и транспортировании каждый блок укладывается на прокладки, которые должны располагаться по вертикали одна над другой между рядами блоков. Подкладки под нижний ряд следует укладывать на плотное, тщательно выровненное основание. Толщина прокладок и подкладок должна быть не менее 30 мм.
Толщина прокладок и подкладок должна быть не менее 30 мм.
Изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов при транспортировании должны быть надежно закреплены с целью предотвращения смещения. Высота штабеля при транспортировании изделий определяется в зависимости от грузоподъемности транспортных средств и допускаемых габаритов погрузки. Погрузку, транспортирование, разгрузку и хранение блоков следует производить с соблюдением мер, исключающих возможность их повреждения.
В компании ГК «БЛОК» можно не только заказать железобетонные фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов, но и проконсультироваться с нашими специалистами, подобрать требуемые конструкции железобетонных изделий. В нашем отделе продаж можно заранее узнать и уточнить цену железобетонных фундаментных блоков и рассчитать общую стоимость заказа. Купить железобетонные изделия фундаментные блоки сплошные ФБС 9.4.6-Т из тяжелого бетона для стен и подвалов и проконсультироваться по общим вопросам покупки и доставки Вы можете, позвонив по телефонам компании ГК «БЛОК»: Санкт-Петербург: (812) 309-22-09, Москва: (495) 646-38-32, Краснодар: (861) 279-36-00. Режим работы компании: Пн-Пт с 9-00 до 18-00. Компания ГК «БЛОК» осуществляет доставку железобетонных фундаментных блоков по всей России прямо до объекта заказчика или на строительную площадку, если позволяет инфраструктура.
Режим работы компании: Пн-Пт с 9-00 до 18-00. Компания ГК «БЛОК» осуществляет доставку железобетонных фундаментных блоков по всей России прямо до объекта заказчика или на строительную площадку, если позволяет инфраструктура.
По вопросам монтажа железобетонных фундаментных блоков обращаться по телефону (812) 309-22-09.
Фундаментные блоки ФБС, ФБС расшифровка, блоки бетонные фундаментные
Главная»Статьи»Фундаментные блоки ФБСФундаментные блоки ФБС – основополагающий материал, который зачастую используется в современном строительстве зданий, благодаря своим качественным характеристикам, простоте монтажа, необычайной прочности и долговечности. ФБС расшифровка звучит как «фундаментные блок стеновой», что и указывает на сферу их применения. Чаще всего, фундаментные блоки используются для сооружения фундаментов самых разнообразных зданий. Благодаря их прочности, такие блоки с большим успехом применяются не только для возведения малоэтажных конструкций, но и при строительстве многоэтажных домов. Кроме того, их часто используют для постройки несущих стен подвалов, технических и цокольных этажей.
Кроме того, их часто используют для постройки несущих стен подвалов, технических и цокольных этажей.
К изделиям такого типа предъявляются самые высокие требования. При производстве фундаментных блоков используется бетон самого высокого класса по прочности. Для увеличения сопротивления ощутимым нагрузкам, довольно часто применяется каркас из железной арматуры. Готовое изделие представляет собой бетонный блок, прямоугольной формы. Стенки блока имеют специальные углубления, которые при монтаже заливаются жидким бетонным раствором. Это придает большую прочность за счет монолитного каркаса, в который превращается сооружение после засыхания бетона. Для удобства транспортировки и укладки такие блоки оснащены петлями из металлической арматуры.
Следует отметить, что для доставки подобных изделий можно воспользоваться услугами самосвала, который может просто и аккуратно высыпать блоки. Фундаментные блоки ФБС настолько прочны, что им не повредит такая процедура. Для монтажа, одним из обязательных факторов, является использование крана или манипулятора, вес конструкций довольно большой, что не позволит обойтись без специальной строительной техники.
Для монтажа, одним из обязательных факторов, является использование крана или манипулятора, вес конструкций довольно большой, что не позволит обойтись без специальной строительной техники.
Блоки бетонные фундаментные выполняют функцию несущего основания. Особенно большими бывают нагрузки при возведении многоэтажных домов. Кроме того, они постоянно подвержены воздействию неблагоприятных факторов из-за того, что находятся в почвенных слоях с повышенной влажностью. Именно поэтому такие изделия должны быть не только максимально прочными, но и устойчивыми к воздействию коррозии и грибка. Не маловажными являются и водонепроницаемость материала, его морозоустойчивость и жаростойкость, способность выдерживать перепады температур. Обладая этими качествами, бетонные блоки являются выгодной альтернативой для строительства высотных и малоэтажных конструкций.
Типы фундаментных блоков и их классификация
Использование готовых бетонных элементов в строительстве – распространённая практика, которая даёт возможность сократить сроки и стоимость работ. При использовании этой технологии не нужно ждать застывания жидкой смеси, после укладки фундамента можно сразу переходить к возведению стен.
При использовании этой технологии не нужно ждать застывания жидкой смеси, после укладки фундамента можно сразу переходить к возведению стен.
ФБС (фундаментные блоки сплошные) используются для сооружения оснований, стен подвалов, несущих опорных конструкций. Изготовить такие элементы можно только в производственных условиях с применением технологии вибропрессования. Компания-производитель «Мир бетона» предлагает купить блоки ФБС с доставкой по Анапе. Заводские изделия изготавливаются из бетонных смесей высоких марок, имеют чёткую геометрию и ровные грани, что обеспечивает лёгкий монтаж.
Классификация блоков ФБС
Требования к блокам определяет ГОСТ 13579-78. Так как эти элементы предназначены для сооружения фундаментов и других ответственных конструкций, для их изготовления применяют тяжёлые, силикатные, керамзитовые бетоны с показателем плотности не меньше 1800 кг/см3. Согласно нормативам, изделия выпускаются в трёх видах:
- ФБС (сплошные). Изготавливаются из бетона высоких марок, имеют наилучшие прочностные характеристики, предназначены для обустройства ленточных сборных фундаментов.
- ФБВ (с вырезом). Аналог сплошных изделий с продольным вырезом для прокладки коммуникаций и проводов.
- ФБП (пустотные). Имеют направленные вниз квадратные пустоты для уменьшения массы и облегчения процесса укладки. Применяются для возведения менее ответственных конструкций, внутренних стен подвалов.
 Изготавливаются из бетона высоких марок, имеют наилучшие прочностные характеристики, предназначены для обустройства ленточных сборных фундаментов.
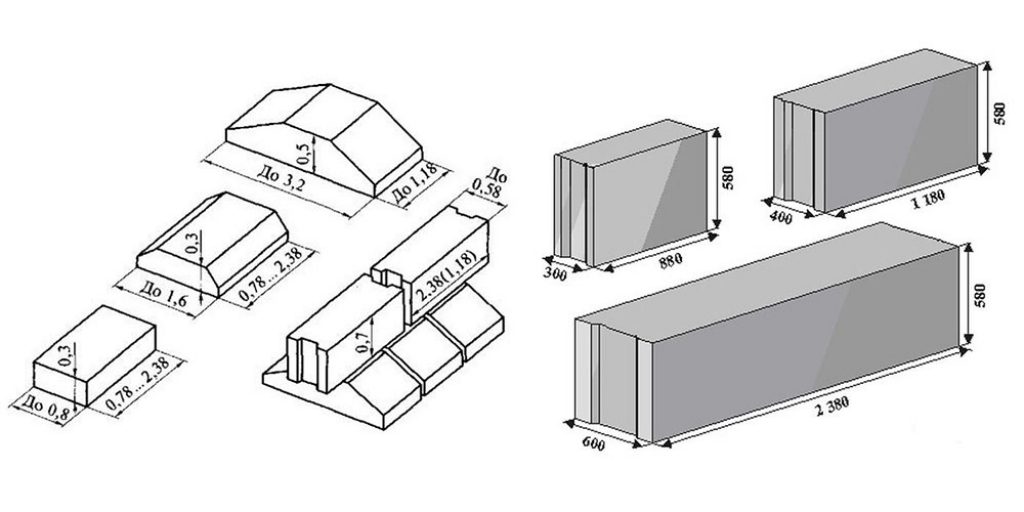
Изготавливаются из бетона высоких марок, имеют наилучшие прочностные характеристики, предназначены для обустройства ленточных сборных фундаментов.Стандартом предусмотрено несколько типоразмеров ФБС, которые соблюдаются всеми производителями (чистые размеры по длине и высоте на 2 см меньше, эта величина предусмотрена на стык между элементами):
- длина – 0,9/1,2/2,4 м;
- высота – 0,3/0,6 м;
- ширина – 0,3/0,4/0,5/0,6 м.
ГОСТ допускает выпуск крупногабаритных нестандартных изделий под заказ.
Расшифровка маркировки
Обычно при возведении строений используется несколько типов ФБС-блоков. Чтобы предотвратить появление ошибки при заказе, необходимо составить план раскладки и рассчитать требуемое количество элементов. В буквенно-цифровой маркировке указана вся необходимая информация:
Чтобы предотвратить появление ошибки при заказе, необходимо составить план раскладки и рассчитать требуемое количество элементов. В буквенно-цифровой маркировке указана вся необходимая информация:
- вид блока;
- размеры в последовательности длина/ширина/высота в дециметрах;
- тип используемого бетона.
Примеры обозначений:
- ФБС-24.3.6 – Т – сплошной, габариты 24*3*6 дм, из тяжёлого бетона;
- ФБП -24.4.6 – С – пустотный, размеры 24*4*6 дм, из силикатного бетона.
В компании «Мир бетона» можно купить разные материалы для индивидуального и промышленного строительства: готовые смеси, растворы, бетонные изделия. Фундаментные блоки 24.3.6 и 24.4.6 – самые востребованные типоразмеры для обустройства ленточных оснований в частном домостроении. Блочные элементы нашего завода имеют высокие показатели влагостойкости, могут применяться на водянистых грунтах. Неограниченный срок службы ФБС позволяет использовать изделия для строительства капитальных сооружений.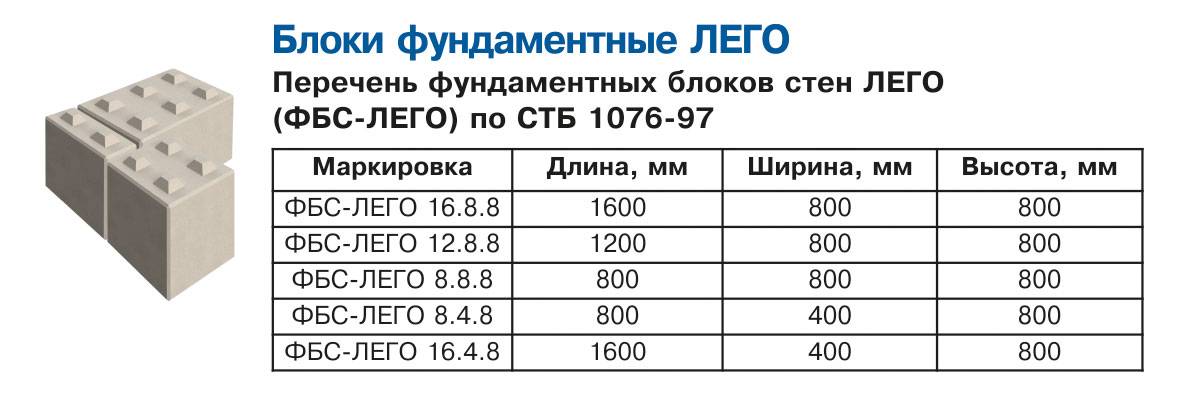
ГОСТ 13579-78
ГОСТ 13579-78
Группа Ж33
MKC 91.080.40
ОКП 58 3500
Дата введения 1979-01-01
1. РАЗРАБОТАН
Центральным научно-исследовательским и проектным институтом типового и экспериментального проектирования жилища (ЦНИИЭП жилища) Госгражданстроя
Всесоюзным научно-исследовательским институтом заводской технологии сборных железобетонных конструкций и изделий (ВНИИжелезобетон) Министерства промышленности строительных материалов СССР
ВНЕСЕН Государственным комитетом по гражданскому строительству и архитектуре при Госстрое СССР
2. УТВЕРЖДЕН И ВВЕДЕН В ДЕЙСТВИЕ Постановлением Государственного комитета Совета Министров СССР по делам строительства от 30.12.77 N 234
3. ВЗАМЕН ГОСТ 13579-68
4. ССЫЛОЧНЫЕ НОРМАТИВНО-ТЕХНИЧЕСКИЕ ДОКУМЕНТЫ
Обозначение НТД, на который дана ссылка | Номер пункта |
ГОСТ 5781-82 | 2. |
ГОСТ 10060.0-95 | 4.3 |
ГОСТ 10060.1-95 | 4.3 |
ГОСТ 10060.2-95 | 4.3 |
ГОСТ 10060.3-95 | 4.3 |
ГОСТ 10060.4-95 | 4.3 |
ГОСТ 10180-90 | 4.1 |
ГОСТ 12730.0-78 | 4.4, 4.5, 4.7 |
ГОСТ 12730.2-78 | 4.7 |
ГОСТ 12730.3-78 | 4.5 |
ГОСТ 12730. | 4.4 |
ГОСТ 13015-2003 | 2.7, 2.12, 3.1, 4.8, 5.1, 5.7 |
ГОСТ 17624-87 | 4.1 |
ГОСТ 18105-86 | 2.2, 2.7, 4.1 |
ГОСТ 21718-84 | 4.7 |
ГОСТ 22690-88 | 4.1 |
СНиП 2.03.01-84 | 2.3 |
СНиП 2.03.11-85 | 2.4 |
СН 165-76 | 2.3, 2.4 |
 9
9 5-84
5-845. ИЗДАНИЕ (октябрь 2005 г.) с Изменением N 1, утвержденным в ноябре 1985 г. (ИУС 3-86)
(ИУС 3-86)
Настоящий стандарт распространяется на блоки, изготовляемые из тяжелого бетона, а также керамзитобетона и плотного силикатного бетона средней плотности (в высушенном до постоянной массы состоянии) не менее 1800 кг/м и предназначаемые для стен подвалов и технических подпольев зданий.
Сплошные блоки допускается применять для фундаментов.
(Измененная редакция, Изм. N 1).
1. ТИПЫ И КОНСТРУКЦИЯ БЛОКОВ
1.1. Блоки подразделяют на три типа:
ФБС — сплошные;
ФБВ — сплошные с вырезом для укладки перемычек и пропуска коммуникаций под потолками подвалов и технических подпольев;
ФБП — пустотные (с открытыми вниз пустотами).
Фундаментные блоки Ростов-на-Дону. Блоки ФБС цены
Компания ДОНСТРОЙМОНОЛИТ предлагает купить фундаментные блоки фбс в Ростове-на-Дону по лояльной стоимости. Высокое качество фундаментных блоков, подтверждено наличием сертификата соответствия и предоставляемой заказчику длительной гарантии на всю линейку продукции.
Форма заказа товаров
Цена блоков ФБС
Мы предлагаем приобрести ЖБИ изделия, соответствующие требованиях ГОСТ, изготовленные из армированного тяжелого бетона. Цена фундаментных блоков зависит от их габаритов. Для удобства потребителей наша компания выполняет доставку купленного товара на строительный участок, причем стоимость такой услуги вполне конкурентоспособна. Ниже приведена таблица цен фундаментных блоков за штуку:
| Название | Ед. изм. | Геом. размеры мм ( L x B x H ) |
Объём, м3 | Вес, т | Цена |
| ФБС 9-3-6т | шт. | 880*300*580 | 0,146 | 0,35 | 600 |
| ФБС 9-4-6т | шт. | 880*400*580 | 0,195 | 0,47 | 700 |
| ФБС 9-5-6т | шт. |
880*500*580 | 0,244 | 0,59 | 800 |
| ФБС 9-6-6т | шт. | 880*600*580 | 0,293 | 0,7 | 900 |
| ФБС 12-3-6т | шт. | 1190*300*580 | 0,203 | 0,487 | 750 |
| ФБС 12-4-6т | шт. | 1190*400*580 | 0,271 | 0,65 | 950 |
| ФБС 12-5-6т | шт. | 1190*500*580 | 0,34 | 0,826 | 1100 |
| ФБС 12-6-6т | шт. | 1190*600*580 | 0,409 | 0,98 | 1250 |
| ФБС 24-3-6т | шт. | 2380*300*580 | 0,406 | 0,97 | 1400 |
| ФБС 24-4-6т | шт.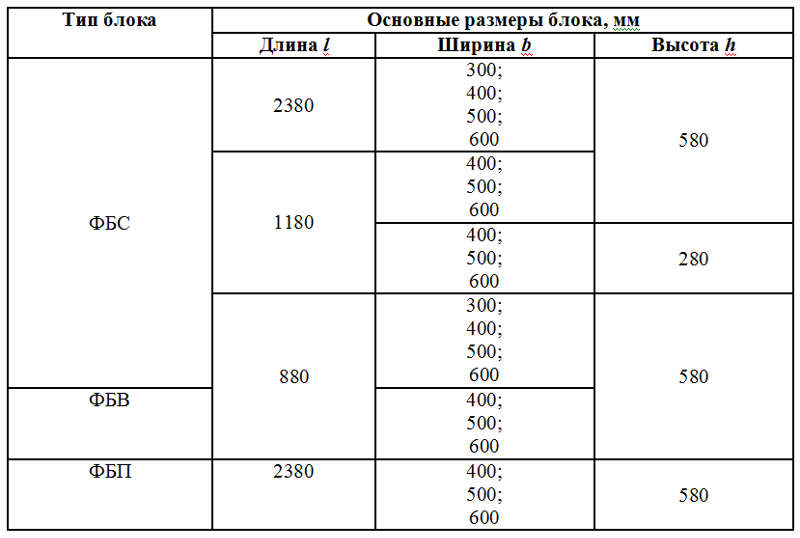 |
2380*400*580 | 0,542 | 1,3 | 1650 |
| ФБС 24-5-6т | шт. | 2380*500*580 | 0,679 | 1,63 | 1900 |
| ФБС 24-6-6т | шт. | 2380*600*580 | 0,817 | 1,96 | 2300 |
* Вся представленная информация, касающаяся стоимости товаров, носит информационный характер, и ни при каких условиях не является публичной офертой, определяемой положениями Статьи 437(2) Гражданского кодекса РФ.
Фундаментные блоки строительные (так расшифровывается аббревиатура ФБС) относятся к наиболее популярным ЖБИ. Их используют при:
- закладке ленточного фундамента;
- установке оснований под рекламные щиты;
- строительстве опорных подушек;
- возведении цокольных этажей;
- постройке подвальных стен и др.

В зависимости от условий эксплуатации (типа почвы, глубины залегания грунтовых вод и т.д.) такие изделия имеют различные коэффициенты прочности и уровни морозоустойчивости. Но в любом случае, для их производства используют бетон класса В 7,5 (М 100), что обеспечивает фундаментным блокам фбс достаточную прочность на сжатие даже при строительстве многоэтажных объектов.
Мы реализуем фундаментные жби, полученные методом виброуплотнения. В результате использования такой технологии в теле фундаментных блоков фбс практически отсутствуют воздушные раковины, что предотвращает разлом изделия при максимально допустимой нагрузке.
В компании ДОНСТРОЙМОНОЛИТ можно купить фундаментные блоки фбс в Ростове, подвергшиеся тепловлажностной обработке в пропарочных камерах при температуре 85-90 градусов. В результате строительные конструкции уже через 12-16 часов приобретают 70% прочность 28 суточного бетона.
Свяжитесь с консультантами нашей компании и вместе подберите подходящие по габаритам и цене фундаментные блоки в Ростове-на-Дону. Одновременно узнайте о стоимости и сроках доставки. Со своей стороны, мы гарантируем, что купленные вами блоки фбс – качественны, долговечны и крепки, а сооружение, стоящее на них, простоит десятилетия.
Одновременно узнайте о стоимости и сроках доставки. Со своей стороны, мы гарантируем, что купленные вами блоки фбс – качественны, долговечны и крепки, а сооружение, стоящее на них, простоит десятилетия.
Форма заказа товаров
Ветров и Температур наверху (FB)
Ветров и Температур наверху (FB)- Ветры и температуры на высотах (FB) — это прогнозы, подготовленные компьютером для определенных мест в континентальной части США и сети местоположений на Аляске и Гавайях, основанные на прогоне модели прогноза Североамериканского мезомасштаба (NAM) [Рис. 1]
- «FDWinds», теперь «FBwinds», производятся как в текстовом, так и в графическом формате.
- Эта информация помогает пилоту:
- Определение наиболее благоприятной высоты на основе ветра и направления полета
- Определение зон возможного обледенения воздушных судов путем регистрации температуры воздуха от + 2 ° C до -20 ° C и температурных инверсий
- Прогнозирование турбулентности путем наблюдения резких изменений направления и скорости ветра на разных высотах
- Есть два основных элемента прогноза ветра и температуры наверху:
- Уровни прогноза
- Выпущено для различных высот в зависимости от местоположения [Рис. 4]
- «FT» указывает уровни ветра и данные температуры
- Группа из четырех цифр показывает направление ветра в десятках градусов, вторые две — скорость ветра в узлах
- Высота до 15000 футов, уровни являются истинной высотой (ссылки на MSL)
- Высота на уровне 18000 футов или выше, уровни являются барометрическими высотами (ссылки на FL)
- Символическая форма прогнозов — DDff + TT, в которой:
- DD — направление ветра
- скорости ветра и
- ТТ температура
- Уровни прогноза
- Данные прогноза ветров и температур на высотах
- Ветер не прогнозируется в пределах 1500 футов от высоты станции
- Ветер прогнозируется истинным, указывается в десятках градусов (две цифры) относительно истинного севера, а скорость ветра указывается в узлах (две цифры).
- Если прогнозируемая скорость менее 5 узлов, кодированная группа — 9900, что означает «легкая и переменная».
- 9 : Ветры легкие и переменные, температура 12 ° C
- Если прогнозируемая скорость больше 100 узлов, 100 вычитается из скорости ветра и 50 добавляется к направлению ветра.
- 731960:
- Шаг 1: 73-50 = 23 или 230
- Шаг 2:19 + 100 = 119
- Результат: 230 @ 119 (температура -60 ° C)
- 731960:
- Если прогнозируется скорость ветра 200 узлов или больше, группа ветра кодируется как 99 узлов.
- 189960: 180 @ 200 + (температура -60 ° C)
- Температура не прогнозируется в пределах 2500 футов над уровнем моря от отметки станции
- В столбце 3000 футов температуры не прогнозируются.
- Группа из шести цифр включает прогноз температуры в градусах Цельсия.
- 192832: последние две цифры показывают температуру 32 ° C, но помните, что выше 24000 ‘отрицательный знак исключается
 4]
4]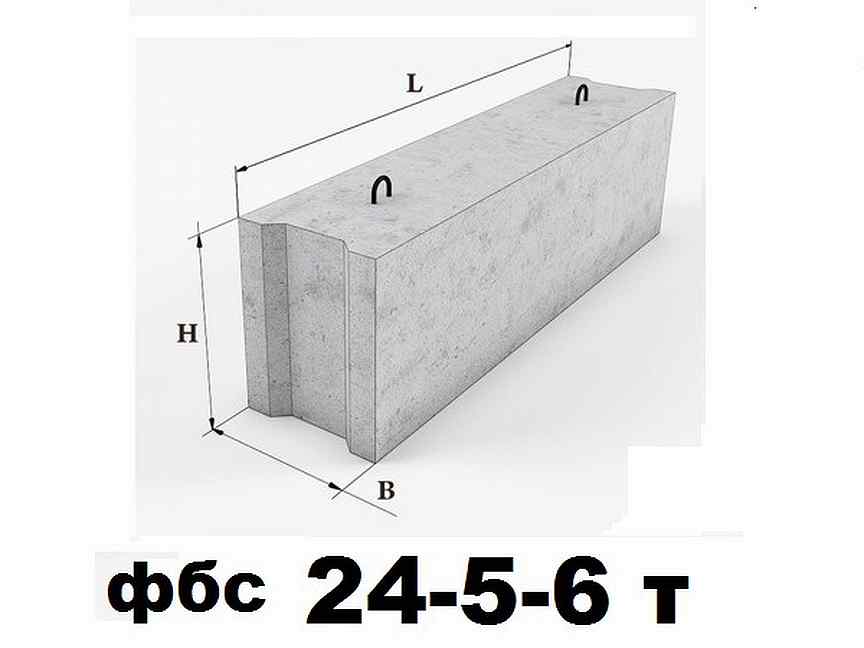
(PDF) Динамические взвешенные алгоритмы декодирования с переворачиванием битов для кодов LDPC
2 IEEE TRANSACTIONS ON COMMUNICATIONS
rule и соответствующий FF. В Разделе IV представлены два расписания обновления весов, алгоритм DWBF
В Разделе IV представлены два расписания обновления весов, алгоритм DWBF
и его производительность, а также некоторые другие однобитовые алгоритмы
. Мы разрабатываем
новое правило FBS и анализируем сложности различных декодеров в
Разделе V. Частота ошибок и поведение сходимости различных многобитовых декодеров BF
, использующих новое правило FBS, также дано
в том же разделе. Наконец, заключительные замечания сделаны в
Разделе VI.
II. ПРЕДПОСЫЛКИ И СМЕЖНЫЕ РАБОТЫ
A. Обозначения и основной алгоритм
Мы обозначим через (N, K) (dv, dc) обычный двоичный код LDPC
C со степенью VN dv и контрольным узлом (CN) степенью dc, т. Е. Cis
нулевое пространство матрицы проверки четности M × N H = [Hmn]
, которая имеет dv1 в каждом столбце и dc1 в каждой строке. Пусть u
будет кодовым словом Cand, предположим, что используется модуляция BPSK
, так что кодовое слово u = (u0, u1, · · ·, uN − 1), ui∈ {0,1},
отображается в биполярную последовательность x = (x0, x1, ···, xN − 1) =
(1 −2u0,1−2u1, ···, 1−2uN − 1) для передачи. Эквивалентный канал передачи основной полосы
Эквивалентный канал передачи основной полосы
представляет собой канал с двоичным входом
с гауссовым выходом, характеризующийся аддитивным нулевым средним
белым гауссовским шумом с двусторонней спектральной плотностью мощности
N0 / 2 Вт / Гц. Пусть y = (y0, y1, …, yN-1) будет последовательностью
значений мягкого канала, полученных на выходе согласованного согласованного фильтра приемника. Последовательность z = (z0, z1, …, zN − 1),
, где zi∈ {0,1}, получается путем принятия жесткого решения для
каждой компоненты y.Пусть ˆ
u = (ˆu0, ˆu1, …, ˆuN − 1) будет
ориентировочная декодированная двоичная последовательность в конце итерации декодирования BF
. Мы вычисляем синдром (контрольную сумму)
вектор s = (s0, s1, …, sM − 1) как s = ˆ
u · HT (mod 2). Далее мы обозначим
n-й VN через vn, набор индексов его
, соединяющих CN, через M (n), а набор индексов VN
, проверенных m-м CN cmby N (m). Индексы CN
Индексы CN
в M (n) определяются ненулевыми элементами n-го столбца
H, тогда как индексы в N (m) — m-й строкой
H.
Общий алгоритм декодирования BF может быть описан с помощью Al-
горифма 1 ниже, который включает в себя три важных параметра:
lmax, максимальное количество итераций, En, FF и B, набор индексов
для ipped бит , или для краткости установленный бит с плавающей точкой (FB).
Этот алгоритм выполняет две основные задачи: 1) вычисление
En (этап 2) и 2) создание набора FB B (этап 3). Наиболее ранние работы
были направлены на улучшение 1) или 2). FF, иногда
, называемый функцией стоимости или функцией инверсии [8], используется в качестве показателя надежности при предварительном решении виртуальной сети.Учитывая значения
FF и правило выбора плавающих битов (FBS), мы выбираем
набор виртуальных сетей и выполняем соответствующие предварительные решения
(биты). Выбор самых ненадежных битов или битов, значения FF
которых превышают пороговое значение, являются двумя наиболее популярными правилами. Для
Для
, первое правило, обычно только один бит сбрасывается, если информация канала с программным значением
используется в FF, что приводит к медленной сходимости
. Напротив, последнее правило часто дает
более высокую скорость сходимости, но, возможно, за счет потери производительности
.Мы кратко рассмотрим известные правила FF и FBS в
следующих параграфах.
Алгоритмы декодирования с переворачиванием битов для алгоритма 1
Установить инициализацию l = 0, ˆ
u = z и вычислить wmn для каждого
n∈ N (m), m = 0,1, …, M — 1.
Шаг 1 Для m = 0,1, …, M −1 вычислить
sm = X
n∈N (m)
ˆunHmn (mod 2). (1)
Если s = 0или l = lmax, остановить декодирование и вывести ˆ
u;
иначе, пусть l = l + 1.
Шаг 2 Для n = 0,1 ,…, N −1, вычислить FF En.
Шаг 3 Используйте FF, полученные на шаге 2, чтобы обновить набор битов fl ipped
B
Шаг 4 Отразить для всех n∈ B и перейти к шагу 1.
B. Функции переключения алгоритмов декодирования BF
Gallager предложил что простая сумма двоичных контрольных сумм
может использоваться в качестве FF [2]
En = −X
m∈M (n)
(1-2 см). (2)
(2) подразумевает, что Значение FF обратно пропорционально надежности решения
бит, поскольку оно является возрастающей функцией от числа
ненулевых контрольных сумм (т.е.е., неудовлетворенные контрольные узлы,
UCN).
Принимая во внимание информацию о канале с мягкими значениями и
, назначая веса контрольной суммы, более поздние модификации FF Галлагера
FF можно описать следующей общей формулой:
En = −α1 · φ (ˆun, yn) −X
m∈M (n)
wmn (1 −2sm), (3)
где α1> 0, φ (ˆun, yn) — показатель надежности, включающий значение канала
и / или битовое решение, и, чтобы быть в соответствии с
(2), wmn ≥0.
Для алгоритма взвешенного BF (WBF) [3] φ (ˆun, yn) = 0
и wmn равно
wmn = min
n′∈N (m)
| yn ′ |, (4 )
Модифицированный алгоритм WBF (MWBF) [4] имеет φ (ˆun, yn) =
| yn |, в то время как улучшенный алгоритм MWBF (IMWBF) [5] использует
тот же φ (ˆun, yn), но заменяет вес контрольной суммы на
wmn = min
n′∈N (m) \ n
| yn ′ | (5)
для веры, переданной от cmto v, должен исключать, что исходный код
произошел от vn.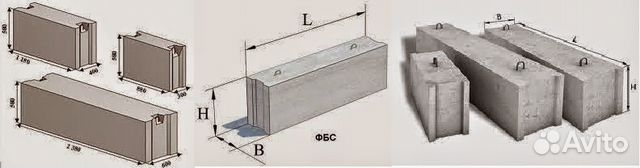 Для алгоритма WBF (RRWBF)
Для алгоритма WBF (RRWBF)
[6], основанного на коэффициенте надежности, φ (ˆun, yn) = 0 и
wmn = 1 / w ′
mn = β | yn |
maxn′∈N (m) | yn ′ | − 1
, (6)
где β — нормализующий коэффициент, гарантирующий, что
Pn∈N (m) w ′
mn = 1.
Алгоритм GDBF Вадаямы и др. [8] применяет метод градиентного спуска
для минимизации
f (ˆ
u) = —
N − 1
X
n = 0
yn (1 −2ˆun) —
M − 1
X
m = 0
(1-2см) (7)
Расшифровка развития крови и иммунной системы во время гемопоэза печени плода человека
Введение
Кровь и иммунная система человека развиваются в раннем эмбриогенезе, и имеют решающее значение для выживания и здоровья организма.На сегодняшний день наше понимание гематопоэтического развития человека основано в основном на мышиных моделях и модельных системах in vitro из-за трудностей доступа к тканям плода человека и экспериментальных манипуляций. В то время как гемопоэтическое развитие сохраняется у позвоночных 1 , важные различия между мышами и людьми были отмечены 2,3 . Например, преобладание гематопоэза печени плода до рождения у мышей 4 и более раннее установление доминирования костного мозга плода (КМ) у людей 5 .Следовательно, исследование тканей человека жизненно важно для понимания молекулярного и клеточного ландшафта развивающейся крови и иммунной системы.
В то время как гемопоэтическое развитие сохраняется у позвоночных 1 , важные различия между мышами и людьми были отмечены 2,3 . Например, преобладание гематопоэза печени плода до рождения у мышей 4 и более раннее установление доминирования костного мозга плода (КМ) у людей 5 .Следовательно, исследование тканей человека жизненно важно для понимания молекулярного и клеточного ландшафта развивающейся крови и иммунной системы.
Последствия гематопоэза плода выходят за пределы жизни in utero до физиологических и патологических состояний у детей и взрослых и имеют отношение к терапевтическому использованию гемопоэтических стволовых клеток (HSC / MPP) и регенеративной медицине. Дальнейшая характеристика эмбриональных HSC / MPP позволит получить информацию о технологии индуцируемых плюрипотентных стволовых клеток и вмешательствах для «перезарядки» пуповинной крови или взрослых HSC / MPP для клинической терапии.Молекулярная карта развивающейся крови и иммунной системы высветит потенциальные механизмы моногенных первичных иммунодефицитов, детских лейкемий 6 и анемий, которые возникают во время жизни плода 7 .
Самые ранние клетки крови и иммунной системы образуются вне эмбриона, возникая из желточного мешка между 2-3 неделями после зачатия (PCW). На 3-4 PCW внутриэмбриональные предшественники из aorta-gonad-mesonephros (AGM) развиваются 8-10 . Предшественники желточного мешка и AGM колонизируют ткани плода, такие как печень, которая остается основным органом кроветворения до середины второго триместра.КМ плода колонизируется около 11 PCW и становится доминантным участком гематопоэза у человека после 20 PCW 8–10 . Иммунные клетки, полученные из желточного мешка, AGM, фетальной печени и костного мозга, засевают периферические ткани, включая нелимфоидные ткани (NLT), где они подвергаются специфическим программам созревания, которые как внутренне детерминированы, так и внешне питаются тканевым микроокружением 11,12 . Систематический комплексный анализ множественных кровеносных и иммунных линий в процессе развития человека ранее не предпринимался.
В этом исследовании мы использовали транскриптомику единичных клеток для картирования молекулярных состояний клеток печени плода человека между 6-18 PCW, когда печень представляет собой преобладающее место гематопоэза плода человека. Мы интегрируем визуализационную массовую цитометрию, проточную цитометрию и клеточную морфологию для проверки клеточных профилей на основе транскриптома. Мы конструируем функциональную организацию развивающейся иммунной сети посредством сравнительного анализа иммунных клеток в печени плода с клетками кожи и почек как репрезентативных нелимфоидных тканей (НЛТ).
Мы интегрируем визуализационную массовую цитометрию, проточную цитометрию и клеточную морфологию для проверки клеточных профилей на основе транскриптома. Мы конструируем функциональную организацию развивающейся иммунной сети посредством сравнительного анализа иммунных клеток в печени плода с клетками кожи и почек как репрезентативных нелимфоидных тканей (НЛТ).
Результаты
Карта транскриптома единичных клеток печени плода
Для исследования крови и развития иммунных клеток в печени плода мы создали суспензии единичных клеток из печени эмбриона и плода между 6 и 18 PCW. Мы FACS-изолированные клетки для капельного (10x Genomics) и индексированного планшетного (Smart-seq2) секвенирования РНК одиночных клеток (scRNA-seq) (рис. 1a-b, таблица с расширенными данными 1, см. Методы). Иммуноокрашивание с помощью CD45 (10x) и CD45 и HLA-DR (Smart-seq2) позволило обогатить клетки в соответствующих воротах (дополнительный рисунок 1a).По возможности, мы анализировали клетки кожи и почек от одних и тех же плодов на ключевой фазе инициации и усиления гематопоэза печени плода между 6-12 PCW. Это позволило провести параллельную оценку топографии крови и иммунных клеток в нелимфоидных тканях (NLT) (рис. 1a-b).
Это позволило провести параллельную оценку топографии крови и иммунных клеток в нелимфоидных тканях (NLT) (рис. 1a-b).
a , Схема обработки ткани и выделения клеток для scRNA-seq. b , Количество клеток печени, кожи и почек плода (n = 15), секвенированных с использованием платформы 10x Genomics, прошедших QC. c , UMAP-визуализация 111,886 фетальных клеток печени с аннотациями. d , средний% каждого типа клеток из c на четырех стадиях развития, скорректированный на основе доли клеток в исходных воротах сортировки (CD45 + / CD45 —), в которых была обнаружена подгруппа. SS2 = Smart-seq2; scRNA-seq = секвенирование РНК одной клетки; HSC / MPP = гематопоэтические стволовые клетки / мультипотентный предшественник; ILC = врожденная лимфоидная клетка; NK = естественный убийца; Нейт-миелоид = миелоид нейтрофилов; Monocyte-DC = моноцит-дендритная клетка; pDC = плазмацитоидная дендритная клетка; DC = дендритная клетка; Моно-мак = моноцит-макрофаг; Моно-NK = моноцит-естественный киллер EI = эритробластический остров; MEMP = мегакариоцит-эритроид-предшественник тучных клеток; PCW = после зачатия недель
Всего мы выполнили scRNA-seq 145 040 клеток печени, из которых 133 756 прошли контроль качества (111 886 клеток с использованием 3′-набора и 21 870 клеток с использованием набора 5 ‘V (D) J; n = 13), 56 800 клеток из кожи с 56 771 проходящим QC (n = 8) и 9 171 клеткой из почек с 9 115 проходящими QC (n = 4) (рисунок 1b и расширенная таблица данных 2).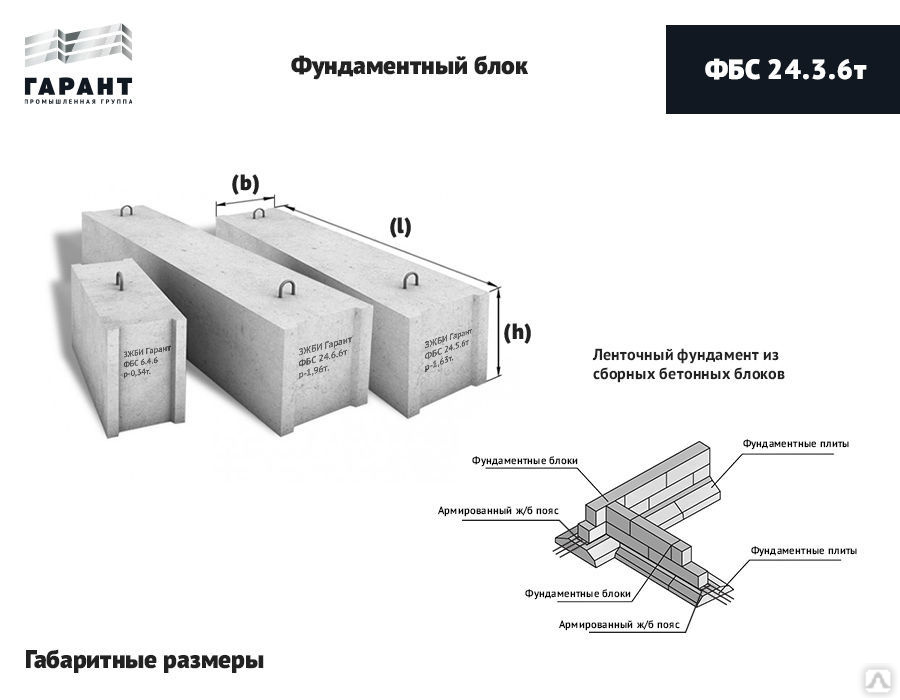 Еще 3571 плодные клетки печени, кожи и почек из 9-12 PCW были профилированы с использованием протокола Smart-seq2 (Таблица 1 с расширенными данными). Мы обнаружили в среднем ~ 3000 генов на клетку с помощью платформы 10x Genomics и ~ 6000 генов с помощью протокола Smart-seq2 (см. Методы). Мы исключили клетки с <200 генами,> 20% экспрессией митохондриальных генов и те, которые идентифицированы как дублеты (см. Методы).
Еще 3571 плодные клетки печени, кожи и почек из 9-12 PCW были профилированы с использованием протокола Smart-seq2 (Таблица 1 с расширенными данными). Мы обнаружили в среднем ~ 3000 генов на клетку с помощью платформы 10x Genomics и ~ 6000 генов с помощью протокола Smart-seq2 (см. Методы). Мы исключили клетки с <200 генами,> 20% экспрессией митохондриальных генов и те, которые идентифицированы как дублеты (см. Методы).
Наш набор данных по печени плода включает 13 кариотипически нормальных плодов: семь женских и шесть мужских (расширенные данные, рис. 1а).Присутствие материнских клеток было исключено с помощью генотипирования считываний мРНК (см. Методы). Мы выполнили кластеризацию Лувена на основе графиков на 111 886 транскриптомах одиночных клеток, созданных 10x 3’kit, и производных маркерных генов, чтобы аннотировать кластеры клеток (см. Методы, рисунок 1c). Согласованность и согласованность кластеров ячеек была дополнительно оценена с помощью метода кластеризации без графа (гауссова смесь) и второго метода кластеризации на основе графа (агломерация с привязкой Уорда) (расширенные данные, рисунок 1b). Выравнивание партий выполняли с использованием Canonical Correlation Analysis 13 и визуализировали с помощью UMAP. В печени плода было идентифицировано 28 основных типов клеток (рис. 1с). Мы аннотировали типы клеток путем перекрестных ссылок на определяющие кластеры транскрипты с опубликованными литературными сообщениями об этих генах и их специфической для клеточного типа экспрессии (см. Методы). Мы применили описательную номенклатуру для этих типов клеток на основе их профиля экспрессии генов.
Выравнивание партий выполняли с использованием Canonical Correlation Analysis 13 и визуализировали с помощью UMAP. В печени плода было идентифицировано 28 основных типов клеток (рис. 1с). Мы аннотировали типы клеток путем перекрестных ссылок на определяющие кластеры транскрипты с опубликованными литературными сообщениями об этих генах и их специфической для клеточного типа экспрессии (см. Методы). Мы применили описательную номенклатуру для этих типов клеток на основе их профиля экспрессии генов.
Чтобы оценить популяции клеток печени плода на разных стадиях развития, мы определили четыре фазы гестации, охватывающие первый и второй триместры: 6-9 PCW (40 194 клеток; n = 4), 9-12 PCW (33055 клеток; n = 4), 12-15 PCW (30 775 ячеек, n = 2) и 15-18 PCW (29 514 ячеек, n = 3) (рисунок 1b) и рассчитали относительную частоту состояний ячеек во время каждой фазы (рисунок 1d).На этих этапах присутствуют все 28 состояний ячеек (Рисунок 1d и Расширенные данные Рисунок 1c), при этом 26/28 состояний ячеек представлены в разреженном наборе данных Smart-seq2 (Расширенные данные Рисунок 1d).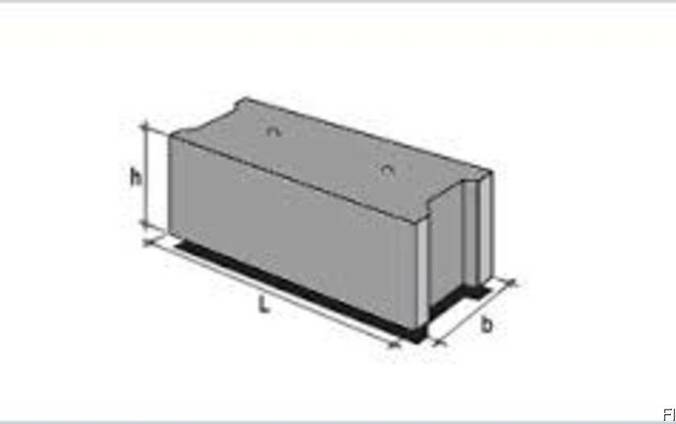 Мы не обнаружили нейтрофилов, базофилов или эозинофилов в печени плода, что согласуется с сообщениями о гранулоцитах, развивающихся позже во время гемопоэза костного мозга плода 14 . Наши результаты согласуются с предыдущими сообщениями о смещении эритроидных клонов в ранней эмбриональной печени и увеличивающейся значимости лимфоидных и миелоидных клонов по мере наступления беременности (рисунок 1d и расширенные данные, рисунок 1 e-f) 5 .
Мы не обнаружили нейтрофилов, базофилов или эозинофилов в печени плода, что согласуется с сообщениями о гранулоцитах, развивающихся позже во время гемопоэза костного мозга плода 14 . Наши результаты согласуются с предыдущими сообщениями о смещении эритроидных клонов в ранней эмбриональной печени и увеличивающейся значимости лимфоидных и миелоидных клонов по мере наступления беременности (рисунок 1d и расширенные данные, рисунок 1 e-f) 5 .
Мы предоставляем полный профиль экспрессии генов, которые, как известно, вызывают первичный иммунодефицит 15 в 28 предполагаемых типах клеток печени плода. Паттерны экспрессии этих генов дадут представление о стадии начала пенетрантности этих мутаций и помогут будущим молекулярным фенотипическим исследованиям этих нарушений (дополнительный рисунок 2).
Проверка маркеров и состояний клеток
Мы получили 150 определяющих кластер маркерных генов (расширенная таблица данных 3) с помощью пакета Seurat (см.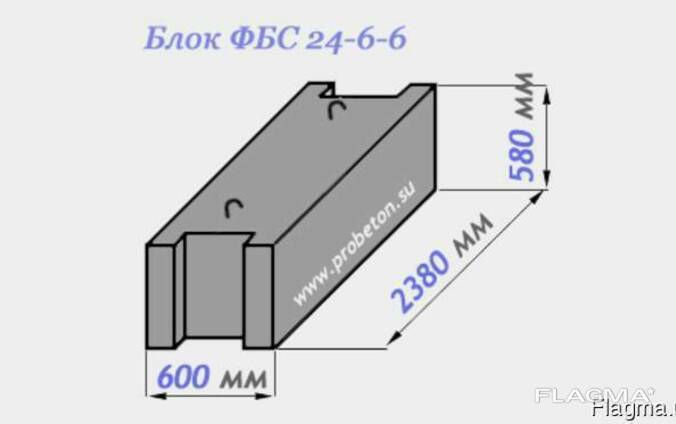 Методы), которые мы вручную усекли до 48 генов (рисунок 2a), где ограниченный набор составляет по-прежнему способен различать все 28 состояний ячеек.Чтобы оценить относительную эффективность списка из 150 генов по сравнению со списком из 48 генов для классификации клеток, мы использовали метод случайного леса с ансамблевым обучением, который дал 92% и 88% средней точности для их соответствующей способности идентифицировать клетки (рис. 2b). Мы выбрали гены, кодирующие поверхностные белки (обозначенные * на рисунке 2a), чтобы разработать панель FACS для выделения клеток, соответствующую кластерам, определенным scRNA-seq (дополнительный рисунок 1b). Мы подтвердили 18 типов клеток печени плода путем массового профилирования транскриптомов из соответствующих ворот FACS (дополнительные рисунки 1b и 3a).Цитоспиновые препараты популяций из этих ворот продемонстрировали морфологию, соответствующую их обозначенной клеточной идентичности, что подтверждает достоверность нашей аннотации и полезность одноклеточной транскриптомики для устранения клеточной гетерогенности (рис.
Методы), которые мы вручную усекли до 48 генов (рисунок 2a), где ограниченный набор составляет по-прежнему способен различать все 28 состояний ячеек.Чтобы оценить относительную эффективность списка из 150 генов по сравнению со списком из 48 генов для классификации клеток, мы использовали метод случайного леса с ансамблевым обучением, который дал 92% и 88% средней точности для их соответствующей способности идентифицировать клетки (рис. 2b). Мы выбрали гены, кодирующие поверхностные белки (обозначенные * на рисунке 2a), чтобы разработать панель FACS для выделения клеток, соответствующую кластерам, определенным scRNA-seq (дополнительный рисунок 1b). Мы подтвердили 18 типов клеток печени плода путем массового профилирования транскриптомов из соответствующих ворот FACS (дополнительные рисунки 1b и 3a).Цитоспиновые препараты популяций из этих ворот продемонстрировали морфологию, соответствующую их обозначенной клеточной идентичности, что подтверждает достоверность нашей аннотации и полезность одноклеточной транскриптомики для устранения клеточной гетерогенности (рис. 2с).
2с).
a , точечная диаграмма выбранных 48 маркерных генов для каждого типа клеток печени из 1c ; * обозначает маркеры, используемые для FACS-выделения клеток. Частота экспрессии генов указывается размером пятна, а уровень — интенсивностью цвета. b , Оценка маркеров клеток печени плода от до с использованием случайного леса для присвоения клеточных меток, фактическое = с использованием всех генов и предсказанное = с использованием выбранных 48 генов. c , окрашенные по Гимзе клетки, полученные цитоспином, выделенные с помощью FACS на основе маркеров с * в a (изображения, репрезентативные для n = 3). d , Наложенные псевдоцветные изображения Hyperion для 8 PCW и 15 PCW печени плода. Крайние левые изображения показаны при 5-кратном увеличении с увеличением вставок справа при 20-кратном увеличении (1 мкм / пиксель).
Затем мы разработали панель маркеров для оценки пространственного распределения эритроидных, тучных клеток, миелоидных и лимфоидных клонов в одном и том же срезе ткани печени плода с использованием визуализирующей массовой цитометрии (рис. 2d). Островки кроветворных клеток вкраплены в синусоиды вокруг агрегатов гепатоцитов по всей печени плода. Макрофаги CD68 + внутри синусоидов были окружены эритроидными клетками GlycophorinA + (рис. 2d). CD1c + DC и CD79a / CD20 + клетки B-линии распределены редко (рис. 2d).Архитектура печени значительно изменилась между 8 и 15 PCW. Рыхлые агрегаты гепатоцитов в печени плода в начале 8 PCW более плотно организованы на 15 PCW, но печеночные дольки с отчетливой центральной веной и портальной триадой четко не наблюдаются. Развитие желчных протоков становится очевидным между 8 и 15 неделями (рис. 2d). Хотя относительная доля гематопоэтических клеток приблизительно соответствовала нашему профилю scRNA-seq, при визуализирующей массовой цитометрии было обнаружено больше гепатоцитов (рис.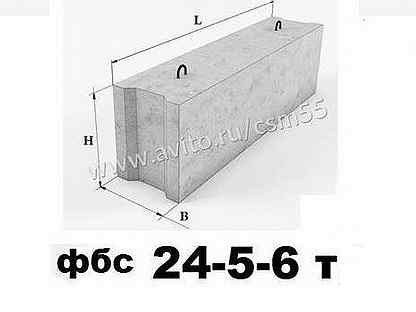 2d и рис. 1c).Это согласуется с опубликованными данными о хрупкости гепатоцитов после выделения ex vivo и высокой экспрессии митохондриальных генов (до 50%) в анализе scRNA-seq взрослых гепатоцитов 16–18 . Проверяя наши наборы данных транскриптома отдельных клеток с помощью нескольких методов, включая проточную цитометрию, морфологию и визуализацию массовой цитометрии для пространственного разрешения, мы предоставляем интегрированную карту гематопоэтических клеток в печени плода.
2d и рис. 1c).Это согласуется с опубликованными данными о хрупкости гепатоцитов после выделения ex vivo и высокой экспрессии митохондриальных генов (до 50%) в анализе scRNA-seq взрослых гепатоцитов 16–18 . Проверяя наши наборы данных транскриптома отдельных клеток с помощью нескольких методов, включая проточную цитометрию, морфологию и визуализацию массовой цитометрии для пространственного разрешения, мы предоставляем интегрированную карту гематопоэтических клеток в печени плода.
Печень плода и гематопоэз NLT
После идентификации ab initio клеток печени плода мы использовали данные в реальном времени из образцов печени на всех стадиях гестации, чтобы вывести траектории развития кроветворных клеток печени плода.Для всех родословных мы использовали три различных вычислительных подхода (силовой граф, диффузионная карта и приблизительная абстракция графа (AGA) с использованием Scanpy) и получили дифференциально экспрессируемые гены в течение псевдовремени с помощью Monocle.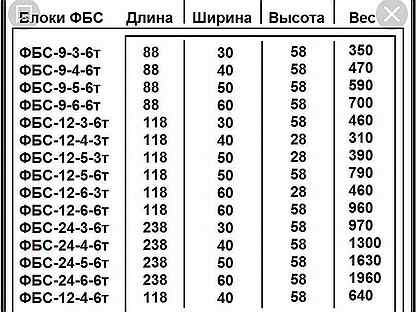 Визуализация графа с принудительным управлением помогла нам идентифицировать три различных соединения с центральным узлом HSC / MPP (рисунок 3a и дополнительное видео 1). Эти связи соответствуют клеткам из следующих родословных; эритроид-мегакариоциты-тучные клетки, В-клетки и врожденные / Т-лимфоидные и миелоидные клоны и обеспечивают всестороннее представление о предполагаемых траекториях гемопоэтической дифференцировки в печени плода человека (рис. 3а).Центральное расположение HSC / MPPs и распределение клеток из соответствующих клонов на Рисунке 3a подтверждают достоверность нашей клеточной аннотации (Рисунок 1c).
Визуализация графа с принудительным управлением помогла нам идентифицировать три различных соединения с центральным узлом HSC / MPP (рисунок 3a и дополнительное видео 1). Эти связи соответствуют клеткам из следующих родословных; эритроид-мегакариоциты-тучные клетки, В-клетки и врожденные / Т-лимфоидные и миелоидные клоны и обеспечивают всестороннее представление о предполагаемых траекториях гемопоэтической дифференцировки в печени плода человека (рис. 3а).Центральное расположение HSC / MPPs и распределение клеток из соответствующих клонов на Рисунке 3a подтверждают достоверность нашей клеточной аннотации (Рисунок 1c).
, Force-ориентированный граф (FDG) визуализация всех кроветворных клеток из 1c . b , FDG-визуализация HSC / MPP печени плода, клонов эритроидных, мегакариоцитов и тучных клеток из a . c , Иммуногистохимическое окрашивание гликофорином А и CD68 печени плода (представитель n = 3).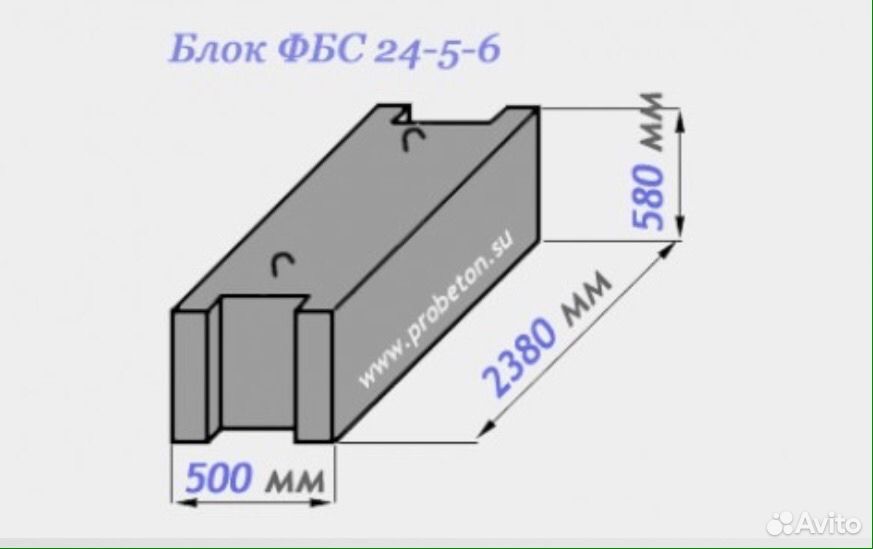 d, Взаимодействия рецептор-лиганд из CellPhoneDB между макрофагами VCAM1 + EI и двумя популяциями эритроидов (ранняя и средняя) со скрипичными графиками, показывающими значение экспрессии генов каждого рецептора и лиганда. Цвета графика скрипки соответствуют предсказанным типам взаимодействующих клеток. и , Тепловая карта 20 лучших экспрессируемых генов в фетальной печени и подмножествах NLT (максимум 20 клеток на подмножество), относительные значения экспрессии от низкого (фиолетовый) до высокого (желтый).
d, Взаимодействия рецептор-лиганд из CellPhoneDB между макрофагами VCAM1 + EI и двумя популяциями эритроидов (ранняя и средняя) со скрипичными графиками, показывающими значение экспрессии генов каждого рецептора и лиганда. Цвета графика скрипки соответствуют предсказанным типам взаимодействующих клеток. и , Тепловая карта 20 лучших экспрессируемых генов в фетальной печени и подмножествах NLT (максимум 20 клеток на подмножество), относительные значения экспрессии от низкого (фиолетовый) до высокого (желтый).
Затем мы сосредоточились на каждой из трех гемопоэтических ветвей, исходящих от узла HSC / MPP в печени плода, и исследовали их связь с гемопоэтическими клетками в фетальном NLT (кожа и почки) (рисунок 3a и дополнительный рисунок 3b).Для сравнения между печенью плода и гематопоэтическими клетками NLT мы использовали шкалу AGA для оценки сходства между тканями и оценивали гены, экспрессирующиеся по-разному, между тканевыми аналогами (см.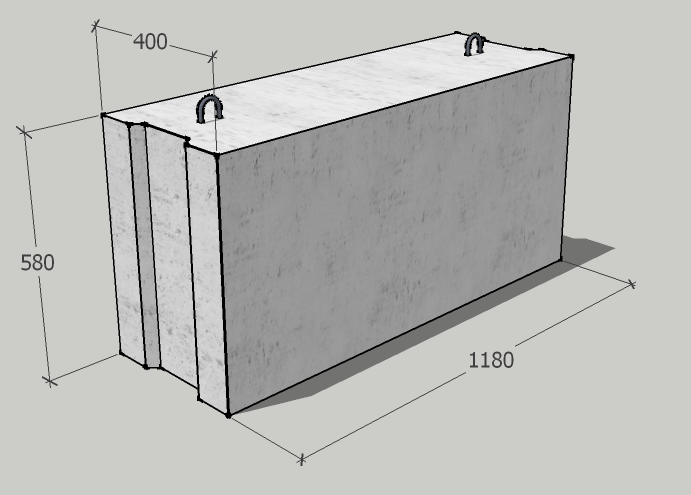 Методы). В печени плода все три подхода к выводу траектории выявляют последовательный результат, предполагая происхождение тучных клеток, эритроидных и мегакариоцитарных клонов от общего предка, расположенного ниже HSC / MPP, предполагаемого предшественника мегакариоцитов-эритроид-тучных клеток (MEMP) (рис. 3b и Рисунок 3а с расширенными данными).Неожиданное наблюдение сцепления тучных клеток с судьбой эритроидов согласуется с наблюдениями за происхождением тучных клеток и базофилов во время гематопоэза мышиного костного мозга и пуповинной крови человека 19,20 . Однако тучные клетки, происходящие из желточного мешка, также наблюдались в коже и соединительных тканях мышей, что указывает на возможное альтернативное происхождение тучных клеток в NLT 19–22 .
Методы). В печени плода все три подхода к выводу траектории выявляют последовательный результат, предполагая происхождение тучных клеток, эритроидных и мегакариоцитарных клонов от общего предка, расположенного ниже HSC / MPP, предполагаемого предшественника мегакариоцитов-эритроид-тучных клеток (MEMP) (рис. 3b и Рисунок 3а с расширенными данными).Неожиданное наблюдение сцепления тучных клеток с судьбой эритроидов согласуется с наблюдениями за происхождением тучных клеток и базофилов во время гематопоэза мышиного костного мозга и пуповинной крови человека 19,20 . Однако тучные клетки, происходящие из желточного мешка, также наблюдались в коже и соединительных тканях мышей, что указывает на возможное альтернативное происхождение тучных клеток в NLT 19–22 .
Мы можем различить три транскрипционно различных стадии развития эритроидов в континууме, характеризующемся ранними, средними и поздними эритробластами с вариабельной экспрессией генов гемоглобина (Рисунок 1c и Расширенные данные Рисунок 3b). Поскольку известно, что эритропоэз печени плода поддерживается за счет адгезивных взаимодействий внутри эритробластных островков (рис. 3c) 23 , мы использовали количественный статистический метод (cellphonedb.org и Vento-Tormo et al., Nature In press ) для прогнозирования клеток. поверхностные взаимодействия рецептор-лиганд между эритробластами и макрофагами эритробластического острова (EI). Наш анализ идентифицировал молекулы, ранее участвовавшие в гематопоэзе, такие как VCAM1, ITGB1, ITGB2, ITGA4 , SIGLEC1, ICAM4 и SPN (рис. 3d) 23–28. С нашими данными с высоким разрешением теперь мы можем точно определить эти молекулы как поддерживающие дифференцировку эритробластов макрофагами EI (рис. 3d).
Поскольку известно, что эритропоэз печени плода поддерживается за счет адгезивных взаимодействий внутри эритробластных островков (рис. 3c) 23 , мы использовали количественный статистический метод (cellphonedb.org и Vento-Tormo et al., Nature In press ) для прогнозирования клеток. поверхностные взаимодействия рецептор-лиганд между эритробластами и макрофагами эритробластического острова (EI). Наш анализ идентифицировал молекулы, ранее участвовавшие в гематопоэзе, такие как VCAM1, ITGB1, ITGB2, ITGA4 , SIGLEC1, ICAM4 и SPN (рис. 3d) 23–28. С нашими данными с высоким разрешением теперь мы можем точно определить эти молекулы как поддерживающие дифференцировку эритробластов макрофагами EI (рис. 3d).
Затем мы оценили гены, которые дифференциально регулируются во время перехода HSC / MPP в эритроидные клетки, мегакариоциты и тучные клетки (расширенные данные, рисунок 3c). Гены, динамически экспрессируемые во время дифференцировки эритроидов, включают известные факторы транскрипции (TF) TAL1 и GATA2 ; BPGM , который связывает гемоглобин и фермент гликолиза. PKLR (расширенные данные, рисунок 3c) 29,30 .
PKLR (расширенные данные, рисунок 3c) 29,30 .
Напротив, дифференцировка мегакариоцитов включает временную регуляцию F2R , PBX1 и MEIS1 , последний, как известно, димеризует и активирует фактор 4 тромбоцитов (PF4) (расширенный рисунок данных 3c) 31 . Интересно, что мы показываем, что HES1 , о котором ранее сообщалось, что он управляет дифференцировкой тучных клеток посредством передачи сигналов Notch, динамически регулируется во время дифференцировки тучных клеток печени плода (Extended Data Figure 3c) 32 .Рассматривая наши данные по NLT кожи и почек, мы показываем, что тучные клетки, мегакариоциты, предполагаемые MEMP и эритроидные клетки также присутствуют в NLT, но HSC / MPP отсутствуют (дополнительный рисунок 3 и расширенные данные рисунка 3d-e). Поскольку HSC / MPP ограничены печенью плода, присутствие MEMP, мегакариоцитов и тучных клеток во всех 3 участках с аналогичными одноклеточными транскриптомами предполагает, что эти клетки могут циркулировать между тканями на этом этапе жизни плода. Мегакариоцитов больше в почках плода, но ранние, средние и поздние эритроидные клетки присутствуют только в коже (расширенные данные, рис. 3d-e).
Мегакариоцитов больше в почках плода, но ранние, средние и поздние эритроидные клетки присутствуют только в коже (расширенные данные, рис. 3d-e).
Мы сравнили высокоэкспрессируемые и дифференциально экспрессируемые гены соответствующих типов клеток в печени, коже и почках плода (рис. 3e). Транскриптом тучных клеток практически идентичен во всех трех тканях, как и транскриптом мегакариоцитов, что согласуется с потенциальной миграцией этих клеток между сайтами (рис. 3e).
Сходство траекторий дифференцировки между клетками в двух участках NLT и в печени плода предполагает одну из двух возможностей: либо (i) происходит локальное созревание предшественников в NLT, либо (ii) клетки на нескольких стадиях дифференцировки выходят из печени плода. и самостоятельно перейти на NLT (расширенные данные, рис. 3e).Отсутствие эритроидных клеток ниже предполагаемого MEMP в почках плода предполагает, что внепеченочный эритропоэз ограничен тканью (Рисунок 3e и Расширенные данные Рисунок 3e).
Чтобы дополнительно оценить, может ли предполагаемый MEMP обладать способностью к расширению в NLT, мы проверили уровни экспрессии генов пролиферации ( MKI67) и клеточного цикла (Seurat CellCycleScoring, см. Методы) (расширенные данные, рисунок 3f). Среди предполагаемых MEMP 50-70% находятся в фазе S и G2 / M в коже и почках плода, в то время как только 25% находятся в фазе S и G2 / M в печени плода (расширенные данные, рисунок 3f).Кроме того, в отличие от печени и почек плода, предполагаемые MEMP в коже также экспрессируют некоторые ранние гены эритробластов, включая MYL4 (Рисунок 3e) 33,34 , что позволяет предположить, что они могут действовать как предшественники in situ в коже. Эти данные предполагают, что на раннем этапе развития кожа в физиологическом состоянии может способствовать эритропоэзу и дополнять эритроидный выброс печени плода.
Лимфоидные клоны в печени плода и NLT
Предыдущие исследования сообщили о присутствии Т- и В-лимфоцитов 35 , NK-клеток 36 и ILC 37 в печени плода человека. Мы наблюдаем две лимфоидные ветви, которые включают клетки линии B и линии NK / T / ILC соответственно (Рисунок 4a и Расширенные данные Рисунок 4a). Интересно, что кластер «ранние лимфоидные / Т-лимфоциты» включает гетерогенные клетки, которые различаются в зависимости от стадии беременности (расширенные данные, рисунок 1c, рисунок 4a и расширенные данные 4b-c).
Мы наблюдаем две лимфоидные ветви, которые включают клетки линии B и линии NK / T / ILC соответственно (Рисунок 4a и Расширенные данные Рисунок 4a). Интересно, что кластер «ранние лимфоидные / Т-лимфоциты» включает гетерогенные клетки, которые различаются в зависимости от стадии беременности (расширенные данные, рисунок 1c, рисунок 4a и расширенные данные 4b-c).
a , FDG-визуализация HSC / MPP печени плода и типов лимфоидных клеток из 1c , показывающая изменения по стадиям развития (6-9 PCW, 12-15 PCW и 15-18 PCW).b , точечная диаграмма, показывающая экспрессию транскриптов V (D) J в типах клеток печени плода (n = 6). Частота экспрессии генов указывается размером пятна, а уровень — интенсивностью цвета. c , Динамически экспрессируемые гены псевдодинамики во время развития В-клеток, факторы транскрипции отмечены *. Окрашенные по Гимзе Pro / Pre B-клетки и B-клетки, полученные цитоспином из 2c (представитель n = 3).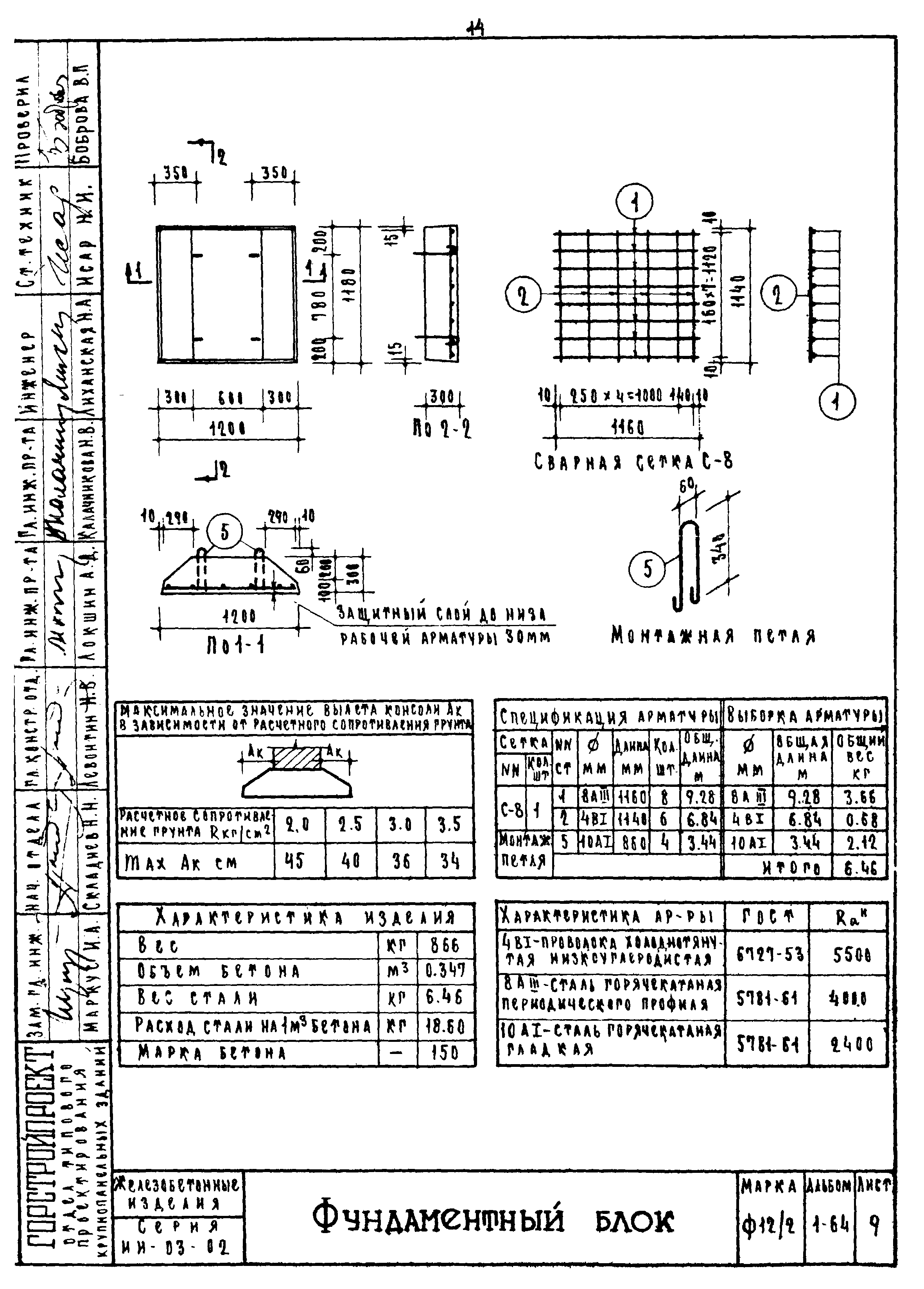 d , FDG-визуализация фетальной печени F1-F10 и соответствующих лимфоидных клеток кожи и почек. и , Графики на скрипке, показывающие экспрессию общих и дифференциально экспрессируемых генов в предшественниках NK и ILC в печени, коже и почках плода F1-F10.
d , FDG-визуализация фетальной печени F1-F10 и соответствующих лимфоидных клеток кожи и почек. и , Графики на скрипке, показывающие экспрессию общих и дифференциально экспрессируемых генов в предшественниках NK и ILC в печени, коже и почках плода F1-F10.
Между 6-9 PCW, до того, как Т-клетки выйдут из тимуса, клетки в этом кластере экспрессируют GATA3 , KLRB1 , CD3D, CD3E, CD7 и JCHAIN (расширенные данные рис. 4b-c), и может относиться к ранее описанным предполагаемым предшественникам тимоцитов печени плода, которые способны генерировать αβT-клетки при совместном культивировании с эпителиальными клетками тимуса 38-40 .После 12 PCW клетки в этом кластере характеризуются экспрессией IL7R, CD2, CD7, CD27 , CD8B с некоторыми клетками, коэкспрессирующими CD4 (расширенные данные 4b-c). Анализ гена рецептора антигена лимфоцитов (из набора 10x Genomics 5 ‘V (D) J) показывает экспрессию дельта-константной цепи Т-клеточного рецептора ( TRDC ) и альфа-константной цепи Т-клеточного рецептора ( TRAC ) клетками из 12-14 PCW, но в клетках из 14-18 PCW отсутствует экспрессия TRDC (рисунок 4b и расширенные данные, рисунок 4c).
В клетках, экспрессирующих TRDC / TRAC , отсутствуют GZMB и PRF1 , цитоплазматические гранулированные продукты, характерные для зрелых CD8 + Т-клеток (рисунок 4b и расширенные данные 4c). Эти результаты соответствуют посеву печени плода клетками γδT и αβT последовательно после их выхода из тимуса после 12 PCW 41 . Это согласуется с предыдущими сообщениями об идентификации Т-клеток только после 18 PCW 40,42 .
Используя карту диффузии и анализ AGA, мы показываем, что NK-клетки (характеризующиеся NCAM1 , EOMES, CD7 , IL2R и CD3E ) и предшественники ILC (характеризующиеся KIT , KLRB3 IL7R , ID2, AHR, RORC , но без IL2R , TBX21 , IL1RL1 ( ST2 )) соединены через лимфоидную ветвь (Рисунок 2a, 4a и Расширенные данные Рисунок 4a).Это открытие согласуется с существующей литературой об общих предшественниках NK и ILC у человека и мыши 43–45 . Интересно, что мы наблюдаем кластер клеток, которые экспрессируют генные модули моноцитов и NK-клеток (кластер Mono-NK), сходные по характеристикам с субпопуляцией моноцитов, ранее идентифицированной в периферической крови взрослого человека 46 .
Интересно, что мы наблюдаем кластер клеток, которые экспрессируют генные модули моноцитов и NK-клеток (кластер Mono-NK), сходные по характеристикам с субпопуляцией моноцитов, ранее идентифицированной в периферической крови взрослого человека 46 .
Клетки B-линии через континуум состояний дифференцировки присутствуют в печени плода (рис. 1c, рис. 3a и 4a).Наиболее примитивный кластер, экспрессирующий CD34 , SPINK2 и транскрипты B-линии IGLL1 и PAX5 , обозначен как «pre pro-B». Кластеры «про-В» и «пре-В» клеток имеют последовательно более высокую экспрессию транскриптов, связанных с В-клетками EBFI , MS4A1 , CD79B, DNTT и генов MHC-II, таких как HLA-DRA, , в то время как экспрессия JCHAIN и LTB снижается (рисунок 2a). Примитивное состояние кластеров «про / пре-В» подтверждается их высоким соотношением ядер к цитоплазме, незрелым хроматином и присутствием ядрышек (рис. 4с).Гены, участвующие в переходе от HSC / MPP к В-клеткам, включают RAC1 , JUN и LDHB (рисунок 4c).
4с).Гены, участвующие в переходе от HSC / MPP к В-клеткам, включают RAC1 , JUN и LDHB (рисунок 4c).
Анализ лимфоидной линии в зависимости от гестационного возраста показывает присутствие пре-B-клеток между 6-9 PCW, но их дифференцировка в зрелые B-клетки в печени плода эффективно завершается только после 9 PCW (рис. 1e и 4a). Мы отметили экспрессию рецептора-лиганда IL10RB-IL10 , которая, как известно, важна для дифференцировки В-клеток 47 , только после 9 PCW в печени плода.Более крупный вопрос о присущих клеткам и факторах тканевого микроокружения, относящихся к гестационному возрасту, поддерживающему дифференцировку В-клеток, требует дальнейших исследований.
Затем мы оценили состав и молекулярные характеристики лимфоидных клеток в фетальном NLT и оценили транскрипционно сходные типы клеток с лимфоидными клетками печени плода, которые дали высокие оценки связности AGA (рисунок 4d и расширенные данные, рисунок 4d). Клетки, похожие на Mono-NK, присутствуют в коже, но имеют более высокую экспрессию генов макрофагов LYVE1 , F13A1, C1QA / B / C и RNASE1 , чем их аналоги в печени плода (расширенные данные, рисунок 4e).Pro-B, пре-B и B-клетки присутствуют в NLT, но HSC / MPP и пре-про-B-клетки отсутствуют (рис. 4d).
Клетки, похожие на Mono-NK, присутствуют в коже, но имеют более высокую экспрессию генов макрофагов LYVE1 , F13A1, C1QA / B / C и RNASE1 , чем их аналоги в печени плода (расширенные данные, рисунок 4e).Pro-B, пре-B и B-клетки присутствуют в NLT, но HSC / MPP и пре-про-B-клетки отсутствуют (рис. 4d).
Предшественники NK, NK-клетки и предшественники ILC, присутствующие в NLT, имеют общую транскрипционную сигнатуру со своими аналогами в печени, однако тканеспецифическая экспрессия хемокинов ( XCLI, CXCL8 ) и генов цитотоксических гранул ( GNLY ) предполагает созревание и адаптацию ткани в коже и почках (рис. 4e). Предшественники ILC в NLT не имеют полных характерных маркеров и TF их зрелых потомков; ILC1, ILC2 и ILC3 (расширенные данные, рис. 4f).Вместе эти находки предполагают, что NLT засеваются предшественниками NK и ILC из печени плода, которые дифференцируют in situ и приобретают тканевый профиль экспрессии генов.
Тканевые сигнатуры в развивающихся миелоидных клетках
На мышах элегантные исследования картирования судеб продемонстрировали, что тканевые макрофаги засеваются желточным мешком и предшественниками фетальной печени 48,49 .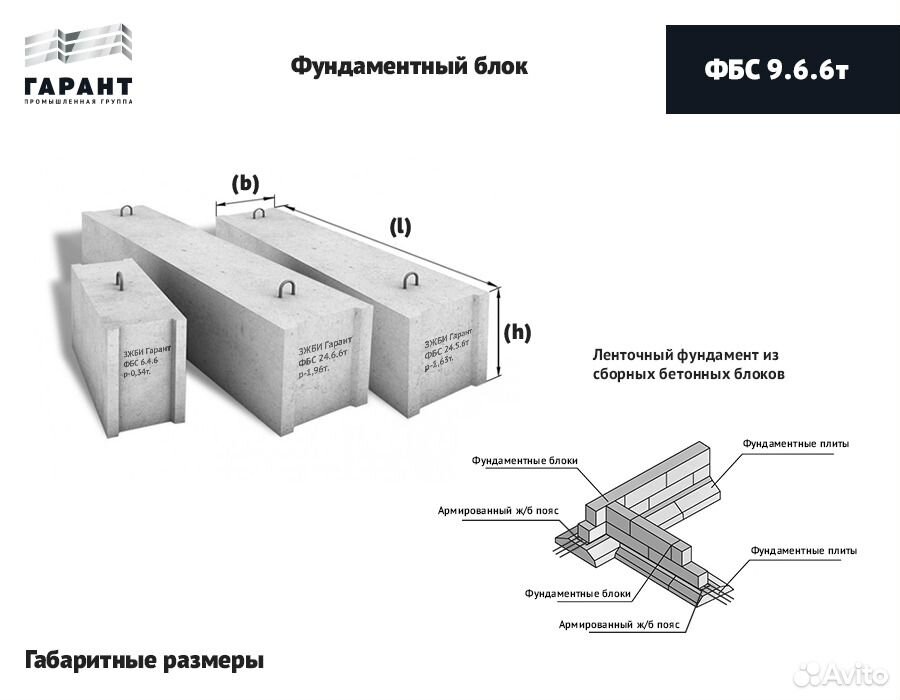 Напротив, дендритные клетки (DC), как полагают, происходят из HSC / MPP, происходящих из костного мозга, через независимую от моноцитов линию 50,51 .Наши данные показывают присутствие миелоидных предшественников, моноцитов, DC и макрофагов уже на 6 PCW в печени плода, а также в NLT, показывая способность HSC / MPP в печени плода также генерировать DC (Рисунок 1c, Расширенные данные Рисунок 1c и Дополнительный рисунок 3b). Неясно, являются ли HSC / MPP-производные DC из печени плода гомеостатически и функционально отличными от BM-HSC / MPP-производных DC.
Напротив, дендритные клетки (DC), как полагают, происходят из HSC / MPP, происходящих из костного мозга, через независимую от моноцитов линию 50,51 .Наши данные показывают присутствие миелоидных предшественников, моноцитов, DC и макрофагов уже на 6 PCW в печени плода, а также в NLT, показывая способность HSC / MPP в печени плода также генерировать DC (Рисунок 1c, Расширенные данные Рисунок 1c и Дополнительный рисунок 3b). Неясно, являются ли HSC / MPP-производные DC из печени плода гомеостатически и функционально отличными от BM-HSC / MPP-производных DC.
Диффузионная карта и анализ AGA миелоидных клеток печени плода выявляют транскрипционно связанные клетки, происходящие из HSC / MPP с увеличивающимся приобретением миелоидных генов (рис. 5a и b и расширенные данные, рис. 5a).Самый ранний миелоидный предшественник нейтрофилов экспрессирует CD34 , SPINK2 и известные миелоидные маркеры, включая GATA2 , ELANE , MPO и LYZ (рис. 2а). Предшественники моноцитов-DC разделяют экспрессию моноцитов и генов DC S100A9 , CD1C и LYZ , но не экспрессируют CD14 . Моноциты экспрессируют CD14 , S100A9 , CD68 , но не CD1C .Обычные DC выражают канонические маркеры для соответствующих подмножеств; CD1C в DC2 и CLEC9A , CADM1 и XCR1 в DC1 (рисунок 2a).
2а). Предшественники моноцитов-DC разделяют экспрессию моноцитов и генов DC S100A9 , CD1C и LYZ , но не экспрессируют CD14 . Моноциты экспрессируют CD14 , S100A9 , CD68 , но не CD1C .Обычные DC выражают канонические маркеры для соответствующих подмножеств; CD1C в DC2 и CLEC9A , CADM1 и XCR1 в DC1 (рисунок 2a).
a , визуализация FDG и b, AGA анализ HSC / MPP фетальной печени и миелоидных клеток из 1c . Цветные облака выделяют линии DC1, DC2 и моноцитов, изображенные на рис. 5c . c , Динамически экспрессируемые гены псевдодинамики во время развития моноцитов, DC1 и DC2, факторы транскрипции, отмеченные *. d , графики скрипки, показывающие значения экспрессии общих и дифференциально экспрессируемых генов в соответствующих миелоидных популяциях в тканях плода (F1-F10).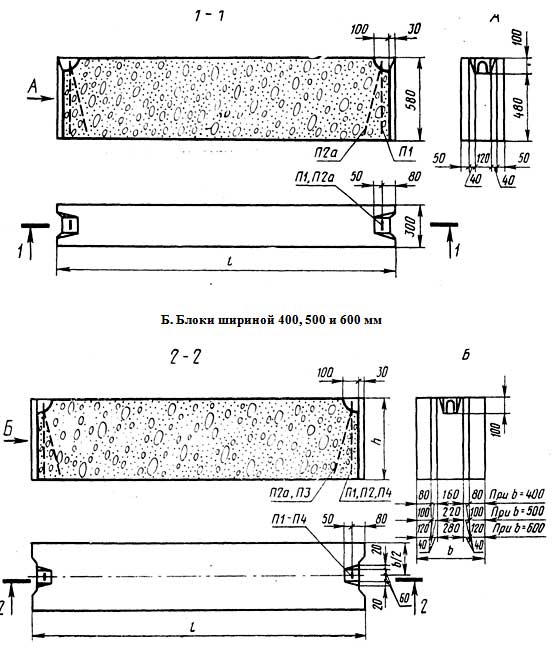
Карта диффузии и анализ AGA предполагает бифуркацию DC1, DC2 и моноцитов от моноцитов-предшественников DC (рис. 5a и 5b, расширенный рисунок 5a). Однако pDCs, по-видимому, ответвляются от двух кластеров клеток: HSC / MPP-ранние про-B-клетки и моноциты-предшественники DC (Рисунок 5a и Расширенные данные Рисунок 5a). Это согласуется с недавними сообщениями об их смешанном лимфоидном и миелоидном происхождении у мышей 19,52–54 .
Анализ AGA показывает только слабые грани (соединительные линии, где толщина определяет транскрипционное сходство) между макрофагами печени плода (клетки Купфера и макрофаги EI) и предшественниками моноцитов-DC (рис. 5b). Это указывает на то, что HSC / MPP фетальной печени являются вероятным источником DC, моноцитов и клеток моно-mac, но что клетки Купфера и макрофаги EI могут возникать из альтернативного источника, такого как предшественники, происходящие из желточного мешка. Было показано, что предшественники желточного мешка вносят вклад в тканевые макрофаги, включая клетки Купфера, у пренатальных мышей 55 , которые могут дожить до взрослого возраста в зависимости от ткани и последующей доступности ниши 56–58 .
Затем мы оценили гены, динамически экспрессируемые во время переходов от HSC / MPP к DC и HSC / MPP к моноцитам-макрофагам независимо (рис. 5c). TF, которые ранее участвовали в дифференцировке DC, такие как GATA2 , IRF8 и BATF3 51 , динамически регулируются в псевдовремя. Мы предполагаем роль дополнительных факторов транскрипции в этих клонах: ATF3 , HES1 и ID3 . В дополнение к каноническим генам, таким как CD14 и FCGR1A / 3A , мы подчеркиваем роль ранее недооцененного члена семейства ядерных рецепторов NR4A2 во время развития моноцитов.
В NLT моноциты, макрофаги и DC2 присутствуют как в коже, так и в почках, pDC и DC1 обнаруживаются только в коже, но мы не можем исключить их присутствие с низкой частотой в почках (дополнительный рисунок 3b). Соответствующие кластеры в NLT и печени плода имеют общее транскрипционное сходство (расширенные данные, рисунок 5b). Тканевые различия в экспрессии генов были наиболее заметны в макрофагах: существует уникальная экспрессия VCAM1 и F13A1 в макрофагах печени 26 и коже 59 соответственно, что напоминает профиль взрослых макрофагов в этих соответствующих тканях (рис. 5г).
5г).
NLT pDC и cDC экспрессируют маркеры созревания и активации DC по сравнению с DC фетальной печени (рисунок 5d и расширенные данные, рисунок 5c). Кожные pDC имели более высокую экспрессию генов MHC-II, CLEC4C (CD303) и NRP1 (CD304), чем pDC фетальной печени, что свидетельствует о более зрелом состоянии (рисунок 5d и расширенные данные, рисунок 5c). DC1 в коже экспрессируют больше S100A4, молекула, участвующая в активации Т-клеток, чем DC1 печени. Экспрессия AOAH , фермента, участвующего в модуляции липополисахаридного ответа, была более специфичной для DC2 печени, тогда как DC2 кожи экспрессировал больше FCER1A и CD83 (фиг. 5d).Эти связанные с тканью генные сигнатуры предполагают приобретение функциональной специализации макрофагами и DC на раннем этапе развития. Появление активированных DCs, несмотря на стерильную среду плода, предполагает активную роль фетальных DCs в обеспечении толерантности, как сообщалось ранее 60 .
Потенциал дифференцировки HSC / MPP при беременности
Наше наблюдение центрального кластера HSC / MPP, из которого излучаются самые ранние коммитированные клетки, согласуется с недавними наблюдениями анализа scRNA-seq у постнатальных мышей и людей 19, 61,62 (Рисунок 6a, Рисунок 6a-c с расширенными данными).Гены, динамически регулируемые во время перехода HSC / MPP к ранним предшественникам, включают узнаваемые TF и рецепторы, участвующие во время гематопоэза, такие как ELANE , LYZ , MPO и AZU1 для нейтрофил-миелоидного клона, LTB JCH, LTB , IL7R и VPERB1 для лимфоидной линии и GATA1 , GATA2 , EPOR и FCER1 для линии мегакариоцит-эритроид-тучных клеток (рис. 6b).
Рис. 6. Потенциал дифференцировки HSC / MPP во время беременности. a , FDG-визуализация HSC / MPP и популяций ранних гематопоэтических предшественников из 1c .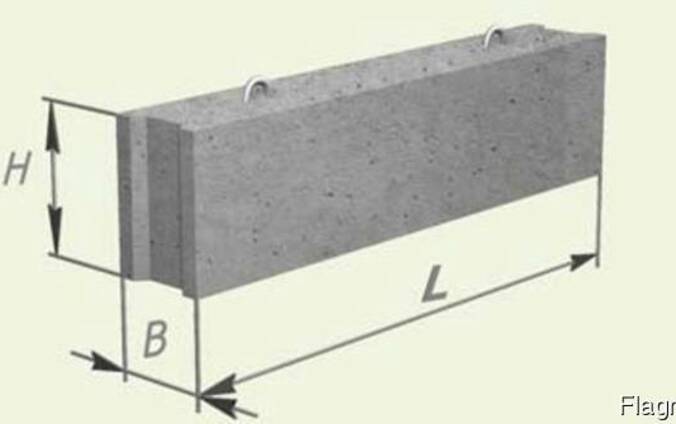 b , Характерные графики динамически экспрессируемых псевдотремовых генов во время перехода HSC / MPP в нейтрофил-миелоидные, MEMP и пре-про В-клетки. Размер точек указывает на уровень экспрессии гена (маленький = низкий; большой = высокий). c , левая панель показывает экспериментальный план для транскриптома одиночных клеток и культуру клеток печени плода из проиллюстрированных ворот FACS, вверху справа показан% колоний, культивируемых из ворот «пула HSC»; 96 клеток, культивированных из каждого образца (n = 5), нижняя правая панель показывает идентичность клеток на основе scRNA-seq для 349 клеток из ворот FACS, соответствующих цвету столбца таблицы (n = 3). d, Процент (медиана +/- SD) HSC, нейтрофил-миелоидных предшественников, MEMP и пре-про B-клеток в печени плода во время развития (n = 13). e, Графики для скрипки (значения экспрессии) дифференциально экспрессируемых генов между HSC / MPP в печени плода (n = 13), пуповинной крови (n = 8) и костном мозге взрослого человека (n = 8).
b , Характерные графики динамически экспрессируемых псевдотремовых генов во время перехода HSC / MPP в нейтрофил-миелоидные, MEMP и пре-про В-клетки. Размер точек указывает на уровень экспрессии гена (маленький = низкий; большой = высокий). c , левая панель показывает экспериментальный план для транскриптома одиночных клеток и культуру клеток печени плода из проиллюстрированных ворот FACS, вверху справа показан% колоний, культивируемых из ворот «пула HSC»; 96 клеток, культивированных из каждого образца (n = 5), нижняя правая панель показывает идентичность клеток на основе scRNA-seq для 349 клеток из ворот FACS, соответствующих цвету столбца таблицы (n = 3). d, Процент (медиана +/- SD) HSC, нейтрофил-миелоидных предшественников, MEMP и пре-про B-клеток в печени плода во время развития (n = 13). e, Графики для скрипки (значения экспрессии) дифференциально экспрессируемых генов между HSC / MPP в печени плода (n = 13), пуповинной крови (n = 8) и костном мозге взрослого человека (n = 8).
Мы предположили, что клеточный состав развивающейся печени плода является результатом локальной модуляции потенциала HSC / MPP. Чтобы проверить эту гипотезу, мы FACS-изолировали отдельные клетки из CD34 + CD38 + , CD34 + CD38 — CD90 + CD45RA — и CD34 + CD38 — CD90 CD45 + FACS и профилировали их как с помощью транскриптомики одиночных клеток на планшете (протокол Smart-seq2), так и анализа потенциала дифференцировки клонов одиночных клеток с использованием установленных систем культивирования 63 (рис. 6c).
Мы использовали SVM, обученную на всем наборе данных о печени плода, чтобы идентифицировать HSC / MPP и предшественников, коммитированных по клону, захваченных анализом Smart-seq2. Сравнивая профили транскриптома с метаданными индекса FACS, используемыми для выделения одиночных клеток (рис. 6c), мы подтверждаем обогащение HSC / MPP (~ 85%) в воротах CD34 + CD38 —, что дополнительно подтверждает нашу аннотацию клеток печени плода. (Рисунок 6c и расширенные данные Рисунок 6d). Большинство клеток в CD34 + CD38 — CD90 — CD45RA + Гейт MLP также содержал HSC / MPP, что согласуется с предыдущими сообщениями о MLP пуповинной крови человека, который транскрипционно наиболее близок к HSC / MPP 64 .
(Рисунок 6c и расширенные данные Рисунок 6d). Большинство клеток в CD34 + CD38 — CD90 — CD45RA + Гейт MLP также содержал HSC / MPP, что согласуется с предыдущими сообщениями о MLP пуповинной крови человека, который транскрипционно наиболее близок к HSC / MPP 64 .
Дифференцированные потомки из культуры единичных клеток также анализировали с помощью проточной цитометрии, чтобы установить потенциал дифференцировки каждой отдельной клетки (расширенные данные, рисунок 6e и дополнительный рисунок 1c). Единичные клетки, культивированные из ворот CD34 + CD38 — (обогащенные HSC / MPP), проявляли способность к мультипотенциальной дифференцировке (расширенные данные, фиг. 6e). Доля колоний, полученных из CD34 + CD38 — одиночных клеток CD45RA — , содержащих NK-клетки, значительно увеличивалась с возрастом гестации, в то время как доля колоний, содержащих эритроидные / мегакариоцитарные клетки, значительно снижалась (расширенные данные, рисунок 6f).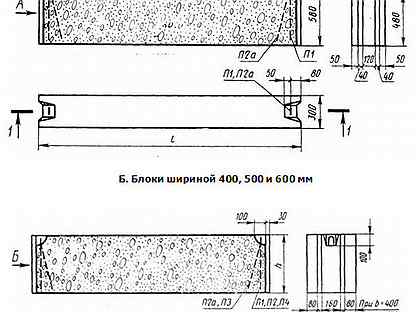
Мы предлагаем этот перекос дифференциации как свойство, присущее HSC / MPP, поскольку жидкие условия культивирования без стромы давали такие же соотношения на выходе (расширенные данные, рисунок 6g). Мы также наблюдали тенденцию к снижению мультипотенциальности с увеличением срока беременности (расширенные данные, рисунок 6e). Эти данные подтверждают гипотезу о дифференциальном внутреннем потенциале HSC / MPP в зависимости от стадии гестации и согласуются с нашими предыдущими наблюдениями об относительном уменьшении количества эритроидных клеток во время развития (рис. 1d).
Мы наблюдали относительное увеличение доли нейтрофил-миелоидных предшественников и снижение количества пре-про-В-клеток до 14 PCW (рис. 6d) с последующим быстрым снижением первых между 14-18 PCW. Это контрастировало с быстрым увеличением относительной частоты HSC / MPP> 14 PCW. Относительное снижение количества ранних предшественников и увеличение HSC / MPP> 14 PCW может быть связано с пролиферативной фазой подготовки к HSC / MPP, засевшей нишу костного мозга, или выходом ранних предшественников (нейтрофил-миелоид, пре-про-B) в BM или даже NLT 65–67 .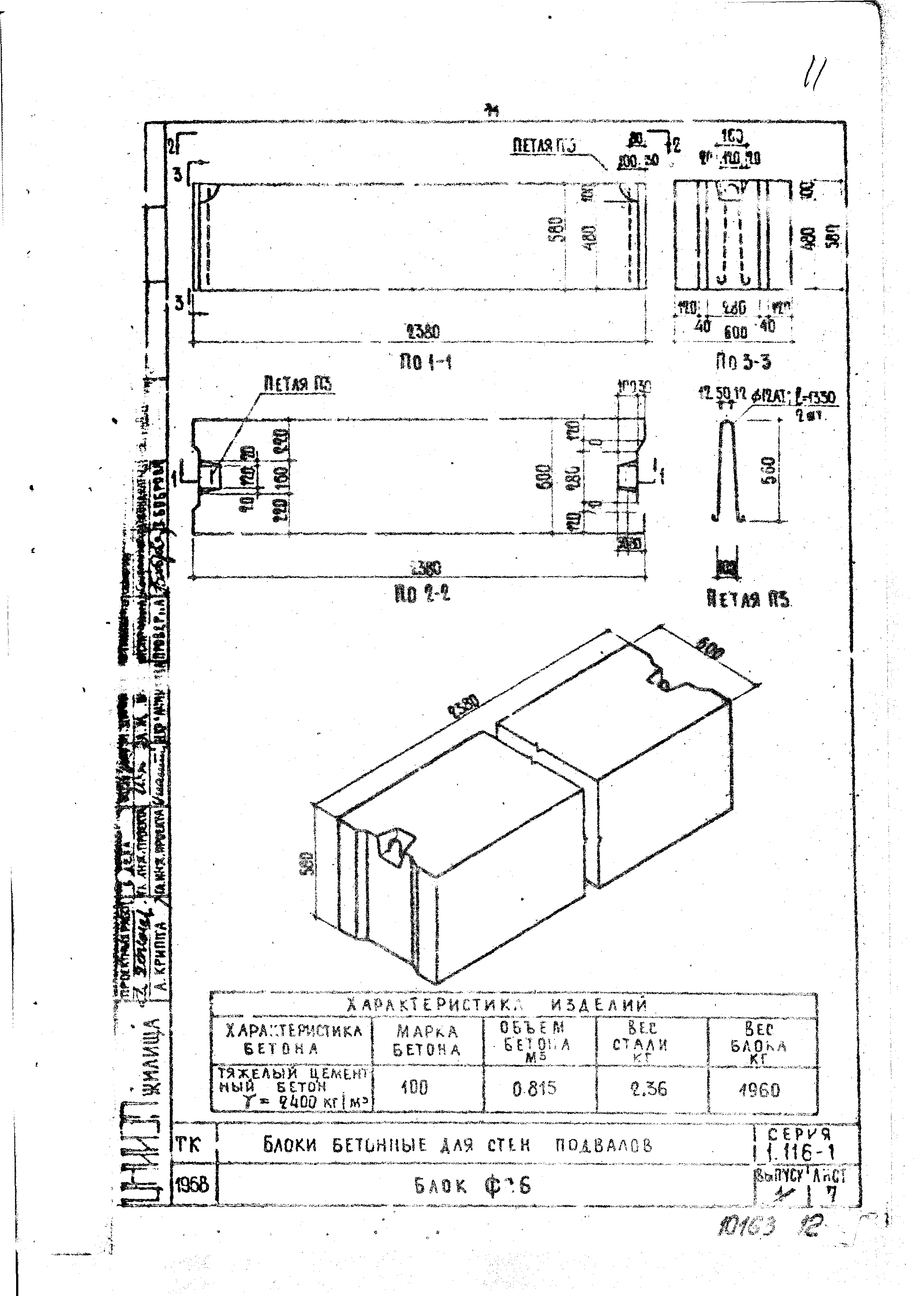 Сравнительный анализ печени и костного мозга плода в будущих исследованиях может дать новое понимание взаимосвязи HSC / MPP и ранних предшественников в этих тканях. Анализ дифференциальной экспрессии генов между HSC / MPP в печени плода, CB и BM взрослого выявляет более высокую экспрессию MKI67 i n HSC / MPP в печени плода, что указывает на повышенный пролиферативный потенциал, как ранее сообщалось у мышей 65–67 , повышенная экспрессия печени -специфические гены ( ALB ), предрасположенность к эритропоэзу ( HBA1) и белок теплового шока ( HSPA1B ), потенциально обеспечивающие тканевую идентичность и поддержание целостности генома и протеома (рисунок 6e и расширенные данные на рисунке 6h).FL HSC / MPP экспрессировали более низкие уровни MHC-I ( HLA-B ), что свидетельствует о снижении антигенпредставляющего потенциала по сравнению с CB и взрослыми BM HSC / MPP (фигура 6e).
Сравнительный анализ печени и костного мозга плода в будущих исследованиях может дать новое понимание взаимосвязи HSC / MPP и ранних предшественников в этих тканях. Анализ дифференциальной экспрессии генов между HSC / MPP в печени плода, CB и BM взрослого выявляет более высокую экспрессию MKI67 i n HSC / MPP в печени плода, что указывает на повышенный пролиферативный потенциал, как ранее сообщалось у мышей 65–67 , повышенная экспрессия печени -специфические гены ( ALB ), предрасположенность к эритропоэзу ( HBA1) и белок теплового шока ( HSPA1B ), потенциально обеспечивающие тканевую идентичность и поддержание целостности генома и протеома (рисунок 6e и расширенные данные на рисунке 6h).FL HSC / MPP экспрессировали более низкие уровни MHC-I ( HLA-B ), что свидетельствует о снижении антигенпредставляющего потенциала по сравнению с CB и взрослыми BM HSC / MPP (фигура 6e).
В совокупности наши результаты демонстрируют, что внутренние изменения в количестве HSC / MPP и потенциале дифференцировки происходят в течение первого и второго триместров плода. Эти изменения, вероятно, будут иметь решающее значение для адаптации кроветворения в печени плода к потребностям развивающегося плода; сначала создание эффективной системы транспорта кислорода, а затем развитие полноценной крови и иммунной системы.
Эти изменения, вероятно, будут иметь решающее значение для адаптации кроветворения в печени плода к потребностям развивающегося плода; сначала создание эффективной системы транспорта кислорода, а затем развитие полноценной крови и иммунной системы.
Обсуждение
Развитие иммунной системы человека in utero остается малоизученным. Предыдущие исследования, как правило, были сосредоточены на одном типе или происхождении иммунных клеток и использовали маркеры взрослых иммунных клеток для идентификации иммунных клеток плода. Используя профилирование одноклеточного транскриптома, мы смогли преодолеть предыдущие предположения о том, что маркеры взрослых достоверно идентифицируют клетки плода и разрешают клеточную гетерогенность в печени плода человека. Анализ печени, кожи и почек плода в течение беременности позволил нам извлечь динамическую временную информацию по тканям, чтобы всесторонне установить состав кроветворных и иммунных клеток во время гематопоэза печени плода.
Мы демонстрируем мощность и валидность классификации клеток на основе транскриптомов путем интеграции данных с помощью нескольких методов: FACS, окрашивание по Гимзе для определения морфологии и визуализация мультиплексных измерений белков на основе массовой цитометрии. Эти методы позволяют экономично и масштабируемо изолировать типы клеток и анализировать их пространственную локализацию.
Наш подход подчеркивает ключевые идеи; тесная связь происхождения тучных клеток с эритромегакариопоэзом, эритропоэз в коже плода как нормальный признак, а не реакция на стресс на заболевание, установление сети DC уже на 6-м PCW, потенциальное двойное миелоидное и лимфоидное происхождение pDC и тканевая адаптация NK , ILC, DC и макрофаги во время развития.Наши результаты показывают модуляцию внутреннего потенциала дифференцировки HSC / MPP в течение срока беременности как функциональный механизм для регулирования гематопоэтического выхода печени плода в течение первого и второго триместров.
В то время как печень плода представляет собой первичный участок эритропоэза плода, наши данные предполагают, что кожа плода также содержит пролиферирующие эритроидные предшественники, которые, вероятно, будут способствовать высокому спросу на развитие крови у плода. Это подтверждается наличием поражений кожи, содержащих экстрамедуллярный гемопоэз, у плодов и новорожденных с тяжелой анемией плода 68 .Хотя процент клеток эритроидного происхождения в коже составляет 1% от такового в печени плода, общая площадь поверхности кожи плода будет поддерживать значительный вклад в эритропоэз, особенно в условиях повышенной потребности в эритропоэзе. Экстрамедуллярное кроветворение в коже может также возникать при врожденной лейкемии, врожденной ЦМВ-инфекции и у взрослых с миелофиброзом, когда наблюдаются папулы, содержащие мегакариоциты 69 . Циркулирующие примитивные эритробласты наблюдались у мышей 70 , а также ранее сообщалось о значительном биогенезе тромбоцитов в постнатальном легком мышей 71 .
Присутствие MEMP, мегакариоцитов и эритроидных клеток на разных стадиях созревания в 6-12 PCW NLT предполагает локальное созревание предшественников или внепеченочный гематопоэз. Приобретение тканеспецифических молекулярных свойств хорошо известно для миелоидных и лимфоидных клеток, включая ILC и Т-клетки 49,72,73 . Это может относиться к функциональной адаптации миелоидных и лимфоидных клеток, но к более общей функции тучных клеток в тканях, на которые транскрипционно не влияет их местное тканевое микроокружение.
Таким образом, наш всеобъемлющий атлас крови плода и иммунных клеток плода человека обеспечивает фундаментальный ресурс для понимания гематопоэза печени плода и развивающейся иммунной системы. Наш справочный набор данных будет неоценимым для исследований педиатрических заболеваний крови и иммунитета, а также использования HSC / MPP для лечения. Наш подход, использующий транскриптомику одиночных клеток для изучения человеческого развития, обеспечивает основу, которая может быть применена для изучения любых временных процессов на протяжении всей жизни человека.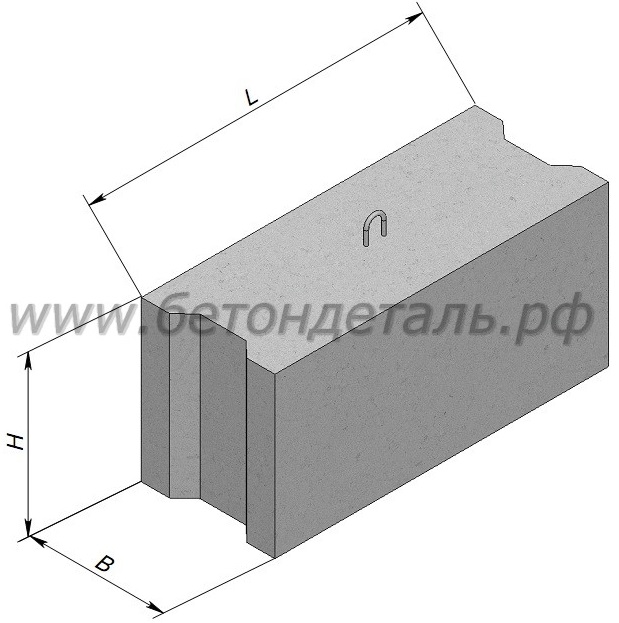
Материалы и методы
Получение тканей
Ткани человеческого плода были получены из Ресурса биологии развития человека, финансируемого MRC / Wellcome Trust (HDBR; http://www.hdbr.org) 74 с соответствующего письменного согласия и одобрения от Объединенного этического комитета Управления здравоохранения NHS Ньюкасла и Северного Тайнсайда (08 / H0906 / 21 + 5). HDBR регулируется Управлением по человеческим тканям Великобритании (HTA; www.hta.gov.uk) и действует в соответствии с соответствующими Кодексами практики HTA.
Обработка тканей
Все ткани обрабатывали сразу после выделения с использованием того же протокола. Ткань переносили в стерильную 10-миллиметровую чашку для культуры ткани 2 и разрезали на сегменты <1 мм 3 перед переносом в коническую пробирку объемом 50 мл. Ткань переваривали 1,6 мг / мл коллагеназы типа IV (Worthington) в RPMI (Sigma-Aldrich) с добавлением 10% (об. / Об.) Термоинактивированной фетальной телячьей сыворотки (Gibco), 100 Ед / мл пенициллина (Sigma-Aldrich), 0,1 мг / мл стрептомицина (Sigma-Aldrich) и 2 мМ L-глутамина (Sigma-Aldrich) в течение 30 минут при 37 ° C с периодическим встряхиванием. Переваренную ткань пропускали через фильтр 100 мкм и клетки собирали центрифугированием (500 г в течение 5 минут при 4 ° C). Клетки обрабатывали 1X буфером для лизиса эритроцитов (eBioscience) в течение 5 минут при комнатной температуре и один раз промывали проточным буфером (PBS, содержащий 5% (об. / Об.) FBS и 2 мМ EDTA) перед подсчетом.
Переваренную ткань пропускали через фильтр 100 мкм и клетки собирали центрифугированием (500 г в течение 5 минут при 4 ° C). Клетки обрабатывали 1X буфером для лизиса эритроцитов (eBioscience) в течение 5 минут при комнатной температуре и один раз промывали проточным буфером (PBS, содержащий 5% (об. / Об.) FBS и 2 мМ EDTA) перед подсчетом.
Определение стадии развития плода и хромосомная оценка
Эмбрионы на сроке до 8 недель после зачатия (PCW) были подвергнуты стадированию с использованием метода стадирования Карнеги 75 .После 8 PCW возраст развития оценивался по измерениям длины стопы и длины от пятки до колена и сравнивался со стандартной диаграммой роста 76 . Кусок кожи или, где это было невозможно, ткань ворсинок хориона собирали из каждого образца для анализа количественной флуоресцентной полимеразной цепной реакции с использованием маркеров половых хромосом и следующих аутосом 13, 15, 16, 18, 21, 22, которые являются наиболее часто встречающимися хромосомными аномалиями.
Проточная цитометрия и FACS для scRNA-seq
Панели антител были разработаны для обеспечения обогащения клеточных фракций для секвенирования и проверки типов клеток.Антитела, используемые для выделения с помощью FACS, перечислены в дополнительной таблице 1. Смесь антител была приготовлена свежей путем добавления 3 мкл каждого антитела в 50 мкл буфера Brilliant Stain Buffer (BD) на ткань. Клетки (≤10 × 10 6 ) ресуспендировали в 50-100 мкл проточного буфера и равный объем смеси антител добавляли к клеткам из каждой ткани. Клетки окрашивали в течение 30 минут на льду, промывали проточным буфером и ресуспендировали при 10 × 10 6 клеток / мл. DAPI (Sigma-Aldrich) добавляли до конечной концентрации 3 мкМ непосредственно перед сортировкой.Сортировку по потоку выполняли на приборе BD FACSAria ™ Fusion с использованием DIVAv8, а данные анализировали с помощью FlowJoV10.4.1. Клетки закрывали для удаления мертвых клеток и дублетов, а затем выделяли для анализа scRNA-seq (10x или Smart-seq2).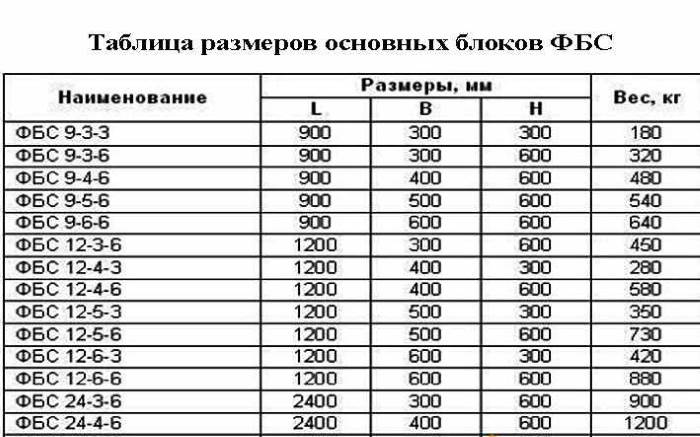 В течение 10 раз клетки сортировали в охлажденные пробирки для FACS, покрытые FBS и предварительно заполненные 500 мкл стерильного PBS. Для Smart-seq2 отдельные клетки сортировали по индексу в 96-луночных планшетах lo-bind (Eppendorf), содержащих 10 мкл лизирующего буфера (TCL (Qiagen) + 1% (об. / Об.) B-меркаптоэтанола) на лунку.
В течение 10 раз клетки сортировали в охлажденные пробирки для FACS, покрытые FBS и предварительно заполненные 500 мкл стерильного PBS. Для Smart-seq2 отдельные клетки сортировали по индексу в 96-луночных планшетах lo-bind (Eppendorf), содержащих 10 мкл лизирующего буфера (TCL (Qiagen) + 1% (об. / Об.) B-меркаптоэтанола) на лунку.
B-клетки также исследовали методом проточной цитометрии согласно Roy et. др., 77 .
Цитоспины и валидация mini-bulk RNA-seq
Замороженные суспендированные клетки из печени плода человека (F15 и F18) размораживали, подсчитывали и разделяли для окрашивания с помощью двух отдельных панелей (см. Дополнительную информацию об антителах в таблице 2). Клетки окрашивали в течение 30 минут на льду с последующим окрашиванием DAPI. FACS выполняли на приборе BD FACSAria ™ Fusion с использованием DIVAv8, а данные анализировали с помощью FlowJov10.4.1. Клетки выделяли в охлажденные пробирки для FACS, покрытые FBS и предварительно заполненные 500 мкл стерильного PBS для цитоспина (500-2000 клеток), или в 1,5 мл микроцентрифужные пробирки, содержащие 20 мкл буфера для лизиса (100 клеток). Окрашивание по Гимзе (Sigma-Aldrich) использовали для определения морфологии отсортированных клеток по цитоспинам. Слайды просматривали с помощью микроскопа Zeiss AxioImager, получали изображения 4 полей из n = 3 образцов с использованием объектива 100x и просматривали с помощью Zen (v2.3).
Окрашивание по Гимзе (Sigma-Aldrich) использовали для определения морфологии отсортированных клеток по цитоспинам. Слайды просматривали с помощью микроскопа Zeiss AxioImager, получали изображения 4 полей из n = 3 образцов с использованием объектива 100x и просматривали с помощью Zen (v2.3).
Культура HSC / MPP
MS5 в логарифмической фазе роста (DSMZ, Германия, пассажи 6-10) высевали в 96-луночные планшеты с плоским дном (поверхность Nunclon delta; Thermo) при плотности 3000 клеток на лунку 24 часов до сортировки.Средой была Myelocult H5100 (Stem Cell Technologies) с добавлением 100 Ед / мл пенициллина и 0,1 мг / мл стрептомицина (Sigma-Aldrich). В день сортировки среду заменяли средой Stem Pro-34 SFM (Life Technologies) с добавлением 100 Ед / мл пенициллина и 0,1 мг / мл стрептомицина, 2 мМ L-глутамина (Sigma-Aldrich), фактора стволовых клеток 100 нг / мл ( Miltenyi), Flt3 20 нг / мл (Miltenyi), TPO 100 нг / мл (Miltenyi), EPO 3 нг / мл (Eprex), IL-6 50 нг / мл (Miltenyi), IL-3 10 нг / мл (Miltenyi), IL-11 50 нг / мл (Miltenyi), GM-CSF 20 нг / мл (Miltenyi), IL-2 10 нг / мл (Miltenyi), IL-7 20 нг / мл (Miltenyi) и липиды (hLDL) 50 нг / мл (Life Technologies) 63 .
Три популяции HSC / MPP и предшественников были выделены из суспензии печени плода (1 свежий F15, 2 замороженных F18, F19). Популяции были идентифицированы по клеткам DAPI —, исключая дублет, как клетки CD3 / CD16 / CD11c / CD14 / CD19 / CD56 —, CD34 + (подробности антител см. В дополнительной таблице 3). Пул HSC / MPP и MLP были обнаружены в 20% клеток с самой низкой экспрессией CD38: пул HSC / MPP был CD90 +/- и CD45RA —, тогда как MLP был CD90 — CD45RA + .Предшественники с наивысшим уровнем экспрессии CD38 20% были отсортированы для сравнения. Отдельные клетки сортировали с использованием BD FACSAria ™ Fusion, работающего с DIVAv8, и сортировали непосредственно на MS5 для культивирования или в 96-луночные планшеты с низким связыванием, содержащие 10 мкл / лунку лизисного буфера для Smart-seq2 scRNAseq. Анализ колоний, полученных из отдельных клеток, выполняли, как описано в Laurenti Lab 63 . Вкратце, колонии собирали в планшеты с 96 U-образным дном, используя фильтр для планшетов, чтобы предотвратить перенос клеток MS5.Клетки окрашивали 50 мкл / лунку вышеуказанного коктейля антител, инкубировали в течение 20 минут в темноте при комнатной температуре, а затем промывали 100 мкл / лунку PBS + 3% FBS. Тип (состав клонов) и размер образовавшихся колоний оценивали с помощью высокопроизводительной проточной цитометрии (BD FACS Symphony). Выход колоний определяли с использованием стратегии стробирования, показанной на Рисунке 6d с расширенными данными. Отдельная клетка определялась как дающая начало колонии, если сумма клеток, обнаруженных в воротах CD45 + и GlyA + , составляла ≥ 30 клеток.Колонии эритроидов были идентифицированы как CD45 — GlyA + ≥ 30 клеток, колонии мегакариоцитов как CD41 + ≥ 30 клеток, миелоидные колонии как [(CD45 + CD14 + ) + (CD45 + CD15 CD15 + + )] ≥ 30 клеток, колонии NK в виде CD45 + CD56 + ≥ 30 клеток. Все данные высокопроизводительной скрининговой проточной цитометрии регистрировались слепым способом, а корреляция между фенотипом колонии и исходной популяцией проводилась только на заключительном этапе.Точный тест Фишера, выполненный в GraphPad (v7.0), был применен к пропорциям каждого типа колонии по этапам, чтобы определить статистическую значимость в потенциале дифференциации линий по мере развития.
Вкратце, колонии собирали в планшеты с 96 U-образным дном, используя фильтр для планшетов, чтобы предотвратить перенос клеток MS5.Клетки окрашивали 50 мкл / лунку вышеуказанного коктейля антител, инкубировали в течение 20 минут в темноте при комнатной температуре, а затем промывали 100 мкл / лунку PBS + 3% FBS. Тип (состав клонов) и размер образовавшихся колоний оценивали с помощью высокопроизводительной проточной цитометрии (BD FACS Symphony). Выход колоний определяли с использованием стратегии стробирования, показанной на Рисунке 6d с расширенными данными. Отдельная клетка определялась как дающая начало колонии, если сумма клеток, обнаруженных в воротах CD45 + и GlyA + , составляла ≥ 30 клеток.Колонии эритроидов были идентифицированы как CD45 — GlyA + ≥ 30 клеток, колонии мегакариоцитов как CD41 + ≥ 30 клеток, миелоидные колонии как [(CD45 + CD14 + ) + (CD45 + CD15 CD15 + + )] ≥ 30 клеток, колонии NK в виде CD45 + CD56 + ≥ 30 клеток. Все данные высокопроизводительной скрининговой проточной цитометрии регистрировались слепым способом, а корреляция между фенотипом колонии и исходной популяцией проводилась только на заключительном этапе.Точный тест Фишера, выполненный в GraphPad (v7.0), был применен к пропорциям каждого типа колонии по этапам, чтобы определить статистическую значимость в потенциале дифференциации линий по мере развития.
Подготовка библиотеки и секвенирование
Для экспериментов по капельной инкапсуляции scRNA-seq 7000 живых, одиночных, CD45 + или CD45 — клеток, выделенных FACS, загружали на каждую дорожку контроллера хрома (10x Genomics) . Библиотеки синтеза, амплификации и секвенирования одной клетки кДНК были созданы с использованием набора реагентов Single Cell 3 ’в соответствии с протоколом 10x Genomics.Библиотеки из восьми загруженных каналов из каждого образца плода были мультиплексированы и секвенированы на Illumina HiSeq 4000. Библиотеки были распределены по восьми дорожкам на проточную кювету и секвенированы с использованием следующих параметров: Read1: 26 циклов, i7: 8 циклов, i5: 0 циклов; Read2: 98 циклов для генерации парных конечных чтений 75 бит / с.
Для экспериментов с scRNA-seq на планшетах использовали слегка модифицированный протокол Smart-seq2, как описано ранее 46 . Всего было проанализировано 45 × 96-луночных планшетов (4320 отдельных клеток) из трех образцов плода.После генерации кДНК готовили библиотеки (384 клетки на библиотеку) с использованием набора Illumina Nextera XT. Наборы Index v2 A, B, C и D использовались для каждой библиотеки для штрих-кодирования каждой ячейки для мультиплексирования. Каждую библиотеку секвенировали (384 клетки) на дорожку при глубине секвенирования 1-2 миллиона считываний на клетку на HiSeq 4000 с использованием химии v4 SBS для создания парных считываний концов 75 п.н.
Для экспериментов mini-bulk RNA-seq каждый клеточный лизат переносили в 96-луночный планшет lo-связывания, затем обрабатывали с использованием того же модифицированного протокола Smart-seq2, как описано выше.После создания кДНК библиотеки были подготовлены с использованием набора Illumina NexteraXT с набором A Index v2 для штрих-кодирования каждой мини-библиотеки для мультиплексирования. Все библиотеки секвенировали на одной дорожке Illumina HiSeq 4000 с использованием химии v4 SBS для создания парных считываний концов 75 пар оснований и с целью достижения глубины 10 миллионов считываний на библиотеку.
Иммуногистохимия
Фиксированные в формалине, залитые парафином блоки печени плода в возрасте 6PCW, 8PCW, 10PCW и 13PCW были получены из HDBR.Каждую из них делали на срезы толщиной 4 мкм на предметные стекла, покрытые APES. Срезы депарафинизировали в течение 5 минут в ксилоле (Fisher Chemical), затем регидратировали с помощью градуированного этанола (99%, 95% и 70%; Fisher Chemical) и промывали проточной водой. Срезы обрабатывали блоком перекиси водорода (1% об. / Об. В воде; Sigma) в течение 10 минут и промывали водопроводной водой перед извлечением антигена. Поиск цитратного антигена использовался для всех разделов. Цитратный буфер, pH 6, использовали при нагревании под давлением для извлечения антигена, а затем слайды помещали в TBS, pH 7.6 за 5 минут до окрашивания. Окрашивание производили с помощью набора Vector Immpress Kit (Vector Laboratories). Срезы сушили и блокировали последовательно 2,5% нормальной лошадиной сывороткой, авидином (Vector Laboratories), а затем биотином (Vector Laboratories) в течение 10 минут каждый и между ними сушили блот. Срезы инкубировали в течение 60 минут с первичным антителом, разведенным в TBS pH 7,6 (подробности антител см. В дополнительной таблице 4). Срезы дважды промывали TBS pH 7,6 в течение 5 минут каждое перед инкубацией в течение 30 минут с вторичным антителом, поставляемым с набором.Предметные стекла дважды промывали TBS pH 7,6 в течение 5 минут каждое и проявляли с использованием пероксидазного хромогена DAB. Срезы контрастировали в гематоксилине Майера в течение 30 секунд, промывали и помещали в водопроводную воду на 30 секунд. Слайды обезвоживали с помощью этанола (от 70% до 99%), а затем помещали в ксилол перед установкой с помощью DPX (Sigma-Aldrich). Срезы получали на микроскопе Nikon Eclipse 80i с использованием NIS-Elements Fv4.
Выравнивание, количественная оценка и контроль качества данных scRNA-seq
Данные секвенирования на основе капель (10x) были выровнены и количественно определены с использованием пакета программного обеспечения Cell Ranger Single-Cell (версия 2.0.2, 10x Genomics Inc) с использованием эталонного генома человека GRCh48 (официальная ссылка Cell Ranger, версия 1.2.0). Данные секвенирования Smart-seq2 были согласованы с STAR (версия 2.5.1b) с использованием индекса STAR и аннотации из той же ссылки, что и данные 10x. Количество считываний, специфичных для генов, было рассчитано с использованием htseq-count (версия 0.10.0). Клетки с менее чем 200 обнаруженными генами, для которых общая экспрессия митохондриальных генов превышала 20%, были удалены. Гены, которые экспрессировались менее чем в 3 клетках, также были удалены.
Обнаружение дублетов машинной классификацией опорных векторов
Исключение дублетов применяли с использованием алгоритма машины опорных векторов (SVM), обученного на данных scRNA-seq и смоделированных дублетах. Данные о дублетах были получены путем случайного выбора пар клеток и сложения их сырых количеств по генам. Каждая пара клеток происходила из одной и той же дорожки, чтобы избежать появления дублетов, которые вряд ли существуют, , то есть . между ячейками, которые были загружены на разных дорожках. SVM была обучена на Python 3.6.3 с использованием SVC (классификатор векторов поддержки) из модуля svm в пакете sklearn (версия 0.19.1). Поиск по сетке применялся во время обучения моделей SVM для оптимизации его гиперпараметров с использованием класса GridSearchCV в sklearn .
Дублетный SVM-классификатор сравнивался с детектором дублетов scrublet и другими настраиваемыми подходами машинного обучения, которые используют случайный лес, k-ближайших соседей и логистическую регрессию.Случайный лес при обучении с оптимизацией параметров поиска по сетке превосходил предсказания данных, близких к 0% дублетов. Логистическая регрессия не соответствовала требованиям, предполагая, что линейные классификаторы могут не подходить для задачи обнаружения дублетов. K-ближайшие соседи, будучи наиболее близкими к scrublet с точки зрения механизма обнаружения, показали высокую зависимость от ручной настройки параметров, что сделало этот метод субъективным.
Критериями для оценки детектора дублетов были: 1) должен обнаруживать очень небольшое количество дублетов или их отсутствие в данных scRNA-seq на планшете, 2) идентифицированные дублеты не должны образовывать компактный отдельный кластер и 3) дублеты должно иметь более высокое количество UMI и генов по сравнению с остальными данными.Оба детектора scrublet и svm удовлетворяли трем критериям, с той разницей, что scrublet предсказал гораздо более низкий процент дублетов (<1%), в то время как детектор svm предсказал скорости 4-6%, которые являются намного ближе к ранее заявленным значениям 78 . Кроме того, мы заметили, что детектор дублетов svm способен удалять больше выбросов, которые, если они присутствуют, искажают карты диффузии.
Кластеризация и последующий анализ данных scRNA-seq
Последующий анализ включал нормализацию данных ( NormalizeData , метод LogNormalize, масштабный коэффициент 10000), масштабирование характеристик данных ( ScaleData ), обнаружение переменных генов (VariableGenes ), pca ( RunPCA , из переменных генов) и кластеризация на основе графа Лувена ( FindClusters , уменьшение размерности данных с помощью PCA, разрешение кластеризации настроено вручную), выполняемые с использованием пакета R Seurat (версия 2.3.4). Идентичность кластерных клеток была присвоена путем ручной аннотации с использованием известных маркерных генов и вычислена дифференциально экспрессируемые гены (DEG) с использованием функции FindAllMarkers в пакете Seurat (критерий суммы рангов Вилкоксона, p — значения, скорректированные для множественного тестирования с поправкой Бонферрони) . Для вычисления DEG были исследованы все гены при условии, что они экспрессировались не менее чем в 25% клеток в любой из двух сравниваемых популяций, и разница экспрессии в логарифмической шкале была не менее 0.25. Ручное аннотирование выполнялось итеративно, что включало проверку предложенных меток клеток с помощью известных маркеров и дальнейшее исследование кластеров, генные сигнатуры которых указывали на дополнительное разнообразие.
Кластеризация и присвоение типа клеток для данных печени плода оценивали с использованием двух дополнительных методов кластеризации: агломеративной кластеризации (со связью Уорда и евклидовым сродством) и гауссовой смесью ( AgglomerativeClustering класс из кластера модуль и GaussianMixture из смеси из модуль в sklearn версия 0.19.1 Python 3.6.3). Согласованность между 3 методами кластеризации была измерена индексом Rand и скорректированной взаимной информацией, реализованной в модуле metrics в пакете sklearn . Индексы rand и скорректированные значения взаимной информации метода кластеризации Лувена по сравнению с методом кластеризации с агломерацией и гауссовой смесью составили 0,84 и 0,83 соответственно.
После завершения аннотации был построен классификатор типов клеток путем обучения SVM на меченых данных scRNA-seq печени плода с поиском по сетке для оптимизации параметров.SVM ранее сравнивался с точки зрения точности и запоминаемости с классификаторами случайного леса и логистической регрессии, обученными на тех же данных. Из трех классификаторов был выбран SVM из-за того, что он показал среднюю точность и запоминаемость 95%. Случайный лес показал 94% точности и запоминания. Логистическая регрессия с регуляризацией l1 и l2 показала 92% точность / отзыв. Более низкая производительность классификаторов логистической регрессии по сравнению с двумя другими была приписана более низкой способности дифференцировать некоторые из ранних предшественников и типы клеток в миелоидном компартменте.Классификатор использовался для автоматического аннотирования наборов данных Smart-seq2, чтобы позволить идентифицировать биологически значимые кластеры и вычислять DEG.
Данные, полученные из кожи и почек плода, были предварительно обработаны, нормализованы, сгруппированы и вручную аннотированы параллельно с данными о печени и с использованием того же конвейера. Данные по коже и почкам были объединены с использованием функции MergeSeurat . Кластеры, характеризующиеся дифференциально экспрессируемыми маркерами иммунных генов, были извлечены из набора данных NLT для последующего сравнительного анализа с иммунными популяциями, полученными из печени.Наборы данных о пуповинной крови и костном мозге взрослого человека были загружены с портала данных Human Cell Atlas (https://preview.data.humancellatlas.org/). Они были обработаны с использованием того же подхода, что и описанный выше, с последующим ручным аннотированием. Выравнивание наборов данных, различающихся технологией секвенирования и пакетной коррекцией, было выполнено с помощью канонического корреляционного анализа ( runCCA , Seurat ).
Методы уменьшения размерности включали tSNE (Seurat, вычисленное с первых 20 компьютеров, быстрое вычисление Barnes-Hut), UMAP (пакет Python UMAP, 5 ближайших соседей, метрика корреляции, минимальное расстояние 0.3, вычислено из первых 20 ПК), FDG (класс ForceAtlas2 из пакета fa2 Python, реализация Barnes-Hut для более быстрых вычислений с тета 0,8, 2000 итераций) и приблизительная абстракция графа на основе разделов (PAGA) ( страниц, в scanpy пакет Python версии 1.2.2). Показатели связности PAGA использовались для количественной оценки сходства типов клеток в тканях и отображались в виде тепловой карты. Траектории развития были выведены путем сравнения графиков FDG, PAGA и диффузионных карт. Анализ траектории включал вычисление карты диффузии ( scanpy tl.diffmap с 20 компонентами), псевдовремя ( scanpy tl.dpt устанавливает самый ранний известный тип клетки как корневой) и изменяемые гены в псевдовремя.
Динамически экспрессируемые гены в псевдовремя
Гены, которые варьируются в псевдовремя, были рассчитаны с использованием функции DifferentialGeneTest в Monocle in R (версия 2.6.4). Это применялось ко всему псевдовременному диапазону, а также к псевдовременным интервалам, специфичным для каждого типа клеток, чтобы избежать ограничения генами, характеризующимися монотонными изменениями по траектории.Перед визуализацией экспрессию генов псевдовременных переменных нормализовали по минимальному и максимальному значениям. Гены псевдовременных переменных были аннотированы на основе участия каждого гена в соответствующих клеточно-специфичных функциональных модулях или основных функциональных путях из MSigDB v6.2, тщательно подобранной базы данных молекулярных сигнатур 79 . Факторы транскрипции были отмечены в наборе данных на основе базы данных прогнозирования факторов транскрипции DBD 80 .
Пиковая экспрессия для каждого гена в течение псевдовремени была рассчитана и сгруппирована в категории «Ранняя», «Средняя» или «Поздняя».В целях наглядности полученные списки генов были минимизированы путем упорядочения их от тех, которые присутствуют в наиболее выбранных функциональных путях, до наименьшего, а также обеспечения охвата в псевдовремени. 30 лучших генов в каждом списке были отображены с помощью функции DoHeatmap пакета Seurat, а полный список генов псевдодействия доступен в интерактивных файлах, сопровождающих карты распространения.
Визуализация посредством анимированного представления принудительно-направленного графа
Анимация FDG была создана с использованием собственной модифицированной версии класса ForceAtlas2 в пакете Python fa2 путем сохранения всех промежуточных состояний (опубликованная версия выводит только конечное состояние и отбрасывает все промежуточные звенья).Координаты FDG на каждой итерации были построены, и полученные графики были собраны в видеоформате mp4 с использованием VideoWriter в пакете Python cv2 (версия 3.3.1).
Выделение и проверка гена дифференциального маркера
Проверка гена дифференциального маркера была выполнена с использованием классификатора случайного леса (класс RandomForestClassifier в ансамбле Модуль пакета sklearn Python, со 100 оценками, gini, критерий разделения , максимальные характеристики 15 и максимальная глубина 10 шпагатов).Чтобы определить, присутствовали ли связанные с тканью вариации транскриптома в эквивалентных иммунных популяциях между печенью, кожей и почками, каждую эквивалентную популяцию брали по очереди и группировали в соответствии с тканью происхождения. Затем была применена функция Seurat FindMarkers попарно между каждым субнабором ткани для создания специфичного для типа клетки списка генов, маркирующих каждую субнабор ткани. Они, в свою очередь, были исследованы на предмет биологической значимости, с репрезентативными маркерами, отображаемыми с использованием функции Сера VlnPlot .
Обработка списка первичных генов иммунодефицита
Информация о заболеваниях и генетическом дефиците была извлечена из Picard et al. 15 и вручную аннотирована, чтобы включить названия символов HGNC для каждого генетического дефекта, связанного с заболеванием, для последующей корреляции с набором данных печени. Заболевания, связанные с ВЗОМТ, были разделены в соответствии с основными категориями Международного союза иммунологических обществ (IUIS) и проанализированы по набору данных scRNA-seq печени. 315 уникальных генов были идентифицированы в наборе данных из 354 врожденных ошибок иммунитета, отмеченных в статье.Для каждой категории заболеваний была создана точечная диаграмма с использованием функции Seurat DotPlot и упорядочена по наивысшей экспрессии для каждого гена и для каждого типа клеток, выделены те типы клеток в каждой категории заболеваний, которые выражают наибольшее количество генов, связанных с генетическим дефектом.
Все графики, представленные в рукописи, были построены с использованием пакета ggplot R или пакета Python matplotlib. Статистическую значимость изменений клеточного состава во время развития исследовали с помощью теста Краскела-Уоллиса с последующим множественным сравнением Данна с использованием GraphPad Prism (v7.0), к частотам популяции на всех этапах.
Анализ CellPhoneDB
Для анализа рецептор-лиганд, показанного на рис. 3d, список согласованных взаимодействий рецептор-лиганд был получен из CellPhoneDB (www.CellPhoneDB.org). Взаимодействия, чьи гены рецептора и лиганда имели ненулевую экспрессию, по крайней мере, в 30% клеток в пределах двух типов клеток, соответственно, считались действительными. Были перечислены рецепторно-лигандные взаимодействия между макрофагами VCAM1 + Erythroblastic Island и двумя популяциями эритроидов (ранняя, средняя).
Секвенирование всего генома и идентификация фетальных клеток
Чтобы идентифицировать материнские клетки, присутствующие в наших данных, мы объединили информацию от секвенирования полногеномной ДНК плода с данными РНК-seq одиночных клеток. Для каждого образца мы измерили частоту аллелей в ДНК плода SNP из проекта «1000 геномов» 81 , попадающих в экзоны с популяционной частотой аллелей, превышающей 1%. Затем мы рассматриваем только те SNP, которые являются гомозиготными по ДНК плода для последующего наблюдения в данных scRNA-seq.SNP считался гомозиготным, если частота его аллелей в ДНК плода была меньше 0,2 или больше 0,8 и имел скорректированное значение p FDR менее 0,01 в соответствии с биномиальным тестом для нулевой гипотезы о том, что частота аллелей в ДНК находился в диапазоне [0,3,0,7].
Частоту аллелей каждого из этих SNP с популяционной частотой аллелей> 1%, которые, как известно, являются гомозиготными по ДНК плода, затем измеряли в каждой клетке в данных scRNA-seq. Любые отклонения от гомозиготности в данных RNA-seq должны быть следствием ошибок секвенирования, редактирования РНК или генотипа клетки, отличного от ДНК плода.Для каждой клетки мы рассчитали общую долю считываний по SNP (выбранным, как описано выше), которые отличаются от генотипа плода. Затем мы предполагаем, что частота отклонений по всему геному из-за ошибок секвенирования и редактирования РНК меньше или равна 2%. Для материнских клеток ожидаемая полногеномная скорость отклонения этих SNP равна половине среднего значения частоты аллелей популяции в опрошенных SNP. Наконец, для каждой клетки мы вычислили апостериорную вероятность того, что эта клетка является плодовой или материнской, принимая биномиальное распределение со скоростью 2% для плодной клетки и половиной средней частоты популяционных аллелей для материнской клетки, и присвоили клетке следующую форму: материнская / плод, если любая из апостериорных вероятностей превышает 99%, в противном случае — неоднозначно.Мы проверили этот метод, используя образцы, для которых были доступны ДНК плода и матери.
Массовая цитометрия «Hyperion» с визуализацией (IMC)
Антитела конъюгировали с металлами с использованием наборов для конъюгации Fluidigm MaxPar и соответствующего метода со следующими модификациями; лантаноиды использовали в концентрации 1,5 мМ и промывали в течение более короткого времени (4 × 5 минут) в буфере W перед элюированием. Сверхчистая вода MilliQ использовалась для любых разбавлений и промывок. Фиксированные в формалине залитые парафином срезы размером 4 мкм, полученные из блоков ткани печени плода 8 и 15 PCW, инкубировали при 60 ° C в течение 1 часа, затем депарафинизировали в ксилоле (Fisher).После регидратации с помощью градуированных спиртов (Fisher) и 5-минутной промывки в воде срезы подвергали тепловому извлечению эпитопа с цитратным буфером (pH-6,0). Затем срезы промывали водой и PBS (Gibco) и блокировали 3% BSA (Sigma-Aldrich) в течение 45 минут. Смесь 8 антител, конъюгированных с металлами, разведенных в 0,5% BSA (см. Дополнительную таблицу 5 для получения подробной информации об антителах, добавляли к срезам для инкубации в течение ночи при 4 ° C в увлажненной камере. Слайды дважды промывали 0.2% Triton X-100 разводили в PBS в течение 8 минут, а затем дважды в PBS в течение 8 минут.
Чтобы контрастировать с ядросодержащими клетками, срезы инкубировали с 312,5 нм (193 Ir) Intercalator-Ir (Fluidigm) в течение 30 минут при комнатной температуре. Затем предметные стекла промывали водой в течение 5 минут и позволяли высохнуть на воздухе при комнатной температуре перед визуализацией на массовом цитометре Hyperion. Используя ожидаемые частоты клеток-мишеней из предыдущих данных флуоресцентной проточной цитометрии, размер области интереса (ROI) был установлен на 2.8 мм на 3,8 мм. Энергия абляции была установлена на 2 дБ при частоте лазера 200 Гц. Каждый сеанс абляции генерировал файл изображения .MCD, содержащий информацию для каждой панорамы и измеренную ROI, посредством чего каждый кусок ткани размером 1 мкм, высвобожденный лазером, анализировался на содержание ионов для каждого канала с помощью Time of Flight. Сегментация отдельных ячеек и выделение признаков выполнялись с помощью CellProfiler (v3.1.5). Ядра были идентифицированы с использованием модуля «IdentifyPrimaryObjects», где входные изображения представляли собой сумму окрашенных ДНК каналов Iridium (191 и 193), построенных модулем «ImageAfterMath».Диапазон диаметров, установленный для идентификации ядер, составлял 4-15 пиксельных единиц. Модуль «ExpandOrShrink» использовался для увеличения площади ядерной сегментации на 3 пикселя для определения площади клетки, а модуль «MeasureObjectIntensity» использовался для определения средней интенсивности для каждого идентифицированного клеточного объекта.
drive.web: мудрые примечания к выпуску
- общие -
В настоящее время поддерживается только американский английский.
- сборка 348 [2021-03-02] -
исправлены некоторые ошибки перетаскивания
улучшенное отслеживание параметров с плавающей запятой
- сборка 347 [2021-02-23] -
исправлена ошибка, из-за которой легенда кнопки тренда fp не отображалась на платформе Windows
- сборка 346 [2021-02-23] -
исправлена ошибка, из-за которой легенды терминала не отображались для оборудования flex2 и flex3
добавлено специальное сообщение об ошибке для случая «Установить IP-адрес» для подсети! = 0 & router == 0
изменен пользовательский интерфейс «Маска подсети» на всплывающее меню вместо текстового поля
добавлена поддержка платформы Apple Silicon
добавлено (скрытое) предпочтение, чтобы заставить полноэкранное окно для режима панели
добавлена поддержка для 'Logic / Most Significant Bit
добавлена поддержка фильтров / кумулятивного скользящего среднего.
- сборка 345 [2020-12-03] -
добавлено предпочтение для увеличения и инверсии масштабирования с помощью колеса прокрутки
добавлена предварительная поддержка E3 v3.09
изменились цены за маркетинг
добавлены криптографические хэши SHA256 для всех платформ
добавлена проверка старых прошивок при импорте
- сборка 344 [2020-10-01] -
исправлена ошибка, из-за которой не разрешались подключения к параметрам, на которые влияют функциональные блоки
добавлена поддержка логики управления / запуска привода.
добавлены предельные значения в диалоговое окно исключения для «Адрес вне допустимого диапазона»
- сборка 343 [2020-08-05] -
исправлена ошибка обработки исключений при проверке криптографии
добавлена поддержка 'Motion Control / Linear Position v3
добавлена поддержка Control / Profiler v2
добавлены дополнительные проверки IP-адресации
- сборка 342 [2020-05-26] -
исправлена ошибка регрессии при обработке имен состояний в редакторе 'Logic / State Machine X-X'
- сборка 341 [2020-05-22] -
исправлена ошибка регрессии при обработке названий рецептов в редакторе 'Утилита / Рецепт'
исправлена ошибка, из-за которой аргументы командной строки не передавались при запуске из файла JAR
обновленное руководство пользователя
- сборка 340 [2020-05-08] -
исправлена ошибка базы данных, при которой некоторые FB 'Parameter' были помечены как устаревшие
- сборка 339 [2020-05-01] -
исправлена ошибка обфускации, из-за которой инструкции не добавлялись в редактор конечного автомата
- сборка 338 [2020-04-29] -
исправлено несколько ошибок, связанных с элементами данных объекта размером более 512 байт
добавлена поддержка для 'Control / State Machine' и 'Control / Sequence List'
добавлена поддержка 'Motion Control / Event Divider'
добавлена поддержка автопреобразования соединения между плавающими и стандартными параметрами
добавлена поддержка «Логического / 8-позиционного селектора битов»
добавлена поддержка OctoBlock 'Parameters / {Analog, Boolean, Enumerated, Hexadecimal, Binary} {Out, In}
добавлена поддержка 'Parameters / {Analog, Boolean, Enumerated, Hexadecimal, Binary} Pass-Through's
исправлена ошибка в «Настройках для доставки», когда явные IP-адреса использовались вместо логического локального адреса
добавлена поддержка "Утилиты / Тревоги"
добавлена поддержка параметров 'Parameters / {Analog, Boolean, Enumerated, Hexadecimal, Binary, Float} {Left, Right}
добавлено нотариальное заверение кода для macOS
- сборка 337 [2019-12-16] -
добавлены проверки для предотвращения переполнения программного пространства
улучшено декодирование некоторых записей журнала
добавлена поддержка «Motion Control / Event Timer v2» и «Motion Control / Event Measurer»
добавлена поддержка «Motion Control / Registration v1» и «Motion Control / Event One Shot».
добавлена поддержка "Floating Point / Calculator" & "Floating Point / Modbus Helper"
- сборка 336 [2019-08-29] -
добавлена поддержка программного обеспечения P2 версии 2.45
- сборка 335 [2019-08-13] -
работал над ошибкой Windows "Shear Affine Transform Text"
обновленная поддержка размеров устройства savvyPanel
исправлена ошибка ограничения адреса в 'Utility / Indirect'
улучшено диалоговое окно «Импорт данных устройства» для несоответствующих случаев
добавлена поддержка Optidrive E3 версии 3.08
добавлена поддержка Optidrive E3 с однофазным выходом
- сборка 334 [2019-05-13] -
добавлена поддержка паролей для плиток ссылок на страницы savvyPanel
- сборка 333 [2019-05-07] -
ПРИМЕЧАНИЕ: обычно не выпускается
добавлена поддержка функциональных блоков «Переключатель с плавающей точкой / 2 входа» и «Переключатель с плавающей точкой / 2 выхода».
добавлена поддержка функциональных блоков 'Floating Point / Track and Hold' и 'Floating Point / Clamp'
добавлена поддержка функциональных блоков 'Floating Point / Moving Average Filter' и 'Floating Point / S Ramp'
- сборка 332 [2019-04-04] -
добавлена поддержка dw24X
добавлена прокрутка всплывающих меню для поддержки маленьких экранов
добавлена поддержка функционального блока 'Floating Point / Comparator'
добавлена поддержка функционального блока Utility / Shared Parameters.
добавлена поддержка программного обеспечения E3 версии 3.07
- сборка 331 [2018-12-11] -
изменена упаковка приложения, поэтому Java больше не требуется в ОС
- сборка 330 [2018-11-08] -
исправлены некоторые ошибки при перетаскивании между малой памятью и большими запоминающими устройствами
исправлена ошибка, из-за которой фантомы E3 могли получать неверные значения параметров, зависящих от модели
убран суффикс "clone" из имен при дублировании предметов
внешние соединения теперь отображаются с другим графическим стилем, чем внутренние соединения
- сборка 329 [2018-08-27] -
добавлена поддержка чисел с плавающей запятой и т. д.добавлена поддержка многострочных заметок и заметок на всех страницах
добавлена поддержка большей постоянной памяти
- сборка 328 [20.04.2018] -
исправлена ошибка, из-за которой устаревшие функциональные блоки могли препятствовать обновлению, клонированию и т. д.
добавлены проверки доступности функционального блока при импорте
добавлена поддержка dw228 (Optidrive E3)
изменены иконки для dw229
- сборка 327 [2018-03-09] -
ПРИМЕЧАНИЕ: обычно не выпускается
исправлена ошибка с плиткой с логотипом
- сборка 326 [2018-02-21] -
исправлена ошибка, которая могла привести к сбою "демонстрационного режима"
- сборка 325 [2018-02-14] -
добавлена поддержка загрузки прошивки JLX
исправлена проблема с упаковкой приложения для 64-битной Windows
- сборка 324 [2018-02-08] -
ПРИМЕЧАНИЕ: обычно не выпускается
обновлена поддержка Internet Demo Devices и Load Demo Phantoms.
добавлена поддержка Java9 в macOS
исправлена ошибка, из-за которой кодировка base64 была нарушена Java9
- сборка 323 [2017-12-14] -
исправлена ошибка, из-за которой время выборки параметра назначения не учитывалось при создании внешних подключений
- сборка 322 [2017-12-06] -
ПРИМЕЧАНИЕ: обычно не выпускается
добавлена поддержка цифровых выходов без затухания на устройствах dw21X
исправлена ошибка, из-за которой импорт рецепта через CSV завершался ошибкой, если рецепт был безымянным
добавлена поддержка записи PIN-кодов 2 и 4 серии PL через последовательный порт
исправлена ошибка, из-за которой типы параметров ODPmomentary не сбрасывались автоматически в фантомах
добавлена поддержка dw224S (SEW LTP-B)
добавлена поддержка dw229 (Generic CANopen Master)
добавлена предварительная поддержка dw228 (Optidrive E3)
исправлена ошибка, из-за которой длинные TLD не принимались в адреса электронной почты
добавлена поддержка новой ливреи
- сборка 321 [2017-06-14] -
добавлена поддержка платформы dwM7
исправлена ошибка, из-за которой внешние соединения терялись из-за изменения Phantom... '
добавлена поддержка плитки для типа параметра ODPmomentary
добавлен «Показать как список» в каталог устройства
добавлен «Сортировать по типу» в каталог устройства
убран фильтр "доступность" из экспорта
исправлены некоторые ошибки, связанные с приводом JL / X
исправлена ошибка, из-за которой сбои связи не всегда указывались в параметрах устройства
- сборка 320 [2016-12-19] -
исправления для электронной коммерции
- сборка 319 [2016-10-28] -
добавлены значки IP66 для Optidrive P2
исправлена ошибка, из-за которой опция -37 или -38 не сообщалась
- сборка 318 [2016-10-28] -
ПРИМЕЧАНИЕ: обычно не выпускается
исправлена ошибка, из-за которой системные пароли savvyPanel не импортировались в dw230s
- сборка 317 [2016-10-27] -
ПРИМЕЧАНИЕ: обычно не выпускается
добавлена поддержка дополнительных размеров dw230
- сборка 316 [2016-09-28] -
добавлена поддержка функционального блока 'Utility / FIFO'
добавлена поддержка функционального блока 'Latches / SR Persistent Latch'
- сборка 315 [2016-06-28] -
добавлена поддержка функционального блока Filters / Sampled Average Filter.
исправлена проблема макета в диалоговом окне «Импорт», которая затрагивала некоторые версии Windows.
исправлена ошибка импорта в фантомы dw230
- сборка 314 [2016-06-28] -
ПРИМЕЧАНИЕ: обычно не выпускается
- сборка 313 [2016-06-16] -
ПРИМЕЧАНИЕ: обычно не выпускается
исправлена ошибка при создании плитки ссылки на страницу на странице сигналов тревоги
улучшено сообщение "Получить подробную информацию" при выходе из пространства пользовательских данных
добавлена поддержка функционального блока Ramps / Master Ramp
в контекстное меню функционального блока добавлено 'Скрыть / Показать метку'
добавлена поддержка функционального блока «Арифметика / Множитель / Делитель (мини)»
добавлена поддержка функционального блока 'Utility / Decay Parameter'
исправлена ошибка, из-за которой бесплатные плитки нельзя было разместить прямо из меню на устройствах без возможности savvyPanel
исправлена ошибка, из-за которой в док могли быть добавлены "фиктивные" параметры
исправлены некоторые проблемы с графикой пользовательского интерфейса
- сборка 312 [2015-11-24] -
обновления для работы с новым оборудованием dw230
обновления для обработки новой версии кремния процессора dwM5
исправлена ошибка в коде сообщения об ошибке
добавлена поддержка функциональных блоков ENC1 Linear Position v2 и ENC2 Linear Position v2.
добавлена предварительная поддержка функционального блока Filters / Batch Average Filter
добавлена предварительная поддержка функционального блока Ramps / Master Ramp
исправлены ошибки в функции импорта для dw230
исправлена ошибка макета в Менеджере купонов, когда к одному модулю применялось много обновлений
исправлена ошибка, из-за которой пользовательские значения для существующих перечислений не копировались из соединения или буфера обмена
добавлена возможность задавать имя страницы панели в диалоговом окне плитки ссылки на панель
добавлена поддержка программного обеспечения Optidrive P2 версии 2.00
добавлены базовые проверки в диалогах «Установить IP-адрес»
- сборка 303 [2015-06-08] -
обновления к руководству пользователя
- сборка 302 [2015-06-04] -
обновления информации о функциональных блоках и руководства пользователя
улучшен интерфейс редактора рецептов
добавлены описания украшений в текст при наведении курсора на устройство
добавлены описания ошибок в текст при наведении курсора мыши
- сборка 301 [2015-04-30] -
версия 4.1
изменен с Java Web Start на программы для конкретных платформ, чтобы избежать ошибок JWS в автономном режиме
исправлено несколько ошибок при добавлении плиток на устройства, у которых нет возможности savvyPanel
исправлена ошибка, из-за которой dw230s опрашивались слишком часто
исправлена ошибка, из-за которой dw230s не всегда мог быть обнаружен через USB
исправлена ошибка, из-за которой данные объекта могли превышать MTU
добавлена поддержка плитки в редакторе программ savvyPanel
исправлены индикаторы прогресса импорта / экспорта
- сборка 281 [2015-03-26] -
исправлена ошибка в функции хеширования системного имени savvyPanel для нелатинских символов
исправлена ошибка, из-за которой удаленные FB не удалялись из существующих выборок
добавлено декодирование для кода события 0x54 (длина сообщения USB Endpoint 2 превышает размер буфера)
модифицированный маршалированный код опроса для USB-устройств для ограничения длины сообщений
добавлены «персонализации» для Sprint Electric и Drive Systems Technology
добавлен пункт меню «Копировать набор обнаружения» в каталог устройства (для использования с savvyPanel)
добавлена поддержка флага dw230 только для чтения (прошивка 0x300B и новее)
добавлена поддержка моделей 600V ODP2
исправлена ошибка, при которой условные операции не отмечались в диалоге калькулятора
исправлена ошибка, из-за которой изменение модели приводило к исключению для последовательных устройств.
изменен код обновления для перехода на версию 4.1
- сборка 280 [2015-01-21] -
добавлена поддержка тайлов 'параметр поверх графики'
ссылки на страницы, параметры и ссылки на устройства теперь можно вставлять и удалять из графических плиток savvyPanel
параметры и устройства теперь можно перетаскивать на графические плитки savvyPanel
исправлена ошибка обработки параметра 'Setpoint' в 'savvyPanel / Dual Monitor' в тайлах savvyPanel
при использовании стрелки «вверх» теперь выбирает конкретную страницу SFD, которая соответствует исходному FB
исправлена ошибка, из-за которой внутренний параметр перекрывал параметр 'Control PID v2' 'Output в SFD
исправлена ошибка, из-за которой фиксированные соединения не отображались в параметре источника для серии PL
обновленная система сборки и т. д.- теперь использует Java 7 (и требует Java 7 или новее)
исправлена ошибка зависания при обработке графики SFD для устройств с последовательным подключением
добавлена поддержка PL-серии версии 6.13
- сборка 279 [2014-10-29] -
исправлена ошибка, из-за которой графические элементы меню выравнивания, перемещения и т. д. не отображались для функциональных блоков устройства
исправлена ошибка в диалоге настройки dw230 для явного обнаружения
добавлены графические элементы панели: резервуар, резервуар, клапан (горизонтальный, клапан (вертикальный), укладчик, сушилка, духовка
реорганизованные меню в редакторе savvyPanel
добавлено "прямое" меню выбора графических плиток в панели savvyPanel
добавлена поддержка перетаскивания устройств в редактор savvyPanel
внесены некоторые изменения в графику savvyPanel (стрелки, ссылки на устройства и т. д.))
реорганизованы категории графических плиток savvyPanel
исправлена ошибка, из-за которой параметры в фантомных устройствах не инициализировались при использовании Java 8
- сборка 278 не использовалась -
- сборка 277 [2014-10-23] -
фиксированные данные параметров для привода E2 для обхода перегруженных идентификаторов параметров
исправлена графика для OptidriveP2 RLY1
исправлены графические заглушки для внутренних параметров функционального блока Ramps / Linear Ramp 2
добавлена статистика использования для получения подробной информации об объектах для фантомов
добавлена поддержка программного обеспечения Optidrive E2 версии 1.20
исправлена ошибка, из-за которой значения по умолчанию в фантомах были неверными для упакованных типов данных
улучшен внешний вид редактора Combinational Logic
добавлены кнопки для копирования настроек масштабирования из входящего соединения или скопированного параметра
добавлены кнопки для копирования пользовательских перечислений из входящего соединения или скопированного параметра
добавлены кнопки для копирования переименования из входящего соединения или скопированного параметра
исправлена ошибка, из-за которой пределы аналогового сеттера не работали правильно для некоторых необычных скейлеров
добавлена поддержка функциональных блоков Control / PID v2 и Control / PI v2
устаревшие функциональные блоки Control / PID v1 и Control / PI v1
добавлена поддержка функционального блока Clamps / Ramped Clamp
добавлена поддержка функционального блока 'Control / Section Control'
добавлена поддержка плиток savvyPanel для работы с устройством (сохранение, импорт, экспорт)
добавлено графическое выравнивание / распределение для выделений
исправлена ошибка обфускации, препятствовавшая созданию фантомов Yaskawa F7
добавлена поддержка функционального блока «Счетчики / Расширенный счетчик»
исправлена ошибка, из-за которой SavvyPanel / Alarm Annunciator не работал для устройств без маршалированного опроса
добавлен графический диалог перемещения для выбора
добавлено графическое "подталкивание" для выделения с помощью ctrl-alt-arrow (1.0) и ctrl-shft-arrow (1 сетка)
исправлена ошибка, из-за которой фантомы могли создавать лишние потоки опроса
исправлена ошибка, приводившая к замедлению работы графики SFD с динамическими элементами
добавлена поддержка плитки savvyPanel «Расширенный счетчик» (для работы с функциональным блоком «Расширенный счетчик»)
добавлена поддержка функционального блока 'Logic / Nybble Splitter'
добавлен фильтр экспорта для PL-серии для параметров без мнемоники при последовательном подключении
- сборка 276 [2014-07-18] -
добавлена автоматическая генерация квитанции по электронной почте для транзакций по кредитной карте
исправлены метки информации о пользователях в фантоме "Получить подробную информацию"
для привода PL исправлена последовательная мнемоника, связанная с функциональным блоком привода 16-битного демультиплексора
добавлена поддержка функционального блока Ramps / Linear Ramp 2
улучшенное представление данных об исключениях InterfaceLock
исправлена ошибка, из-за которой «Загрузить прошивку» и «Загрузить программное обеспечение PL / X» могли не сработать, если было включено автоматическое обнаружение.
исправлена ошибка, из-за которой для транзакций по кредитной карте использовалась неправильная дата истечения срока действия
добавлена поддержка PL-серии версий 6.11 и 6.12
- сборка 275 [2014-04-28] -
добавлена поддержка dw230
исправлена ошибка, из-за которой при вставке параметров в SFD мог отображаться неправильный символ, если был включен параметр «Показать значения параметров».
исправлена ошибка в удалении группы
исправлена ошибка перетаскивания
добавлен диалог подтверждения при применении обновлений
исправлена ошибка, из-за которой пользовательские данные терялись из элементов, перетаскиваемых между устройствами
исправлена ошибка, из-за которой имя системы savvyPanel терялось при модификации фантома
добавлена поддержка функционального блока 'Utility / Indirect'
добавлена поддержка функционального блока Encoder / ENC Float IO
исправлена ошибка, из-за которой PL-серия 6.Программное обеспечение XX не было доступно для серийных фантомов
Исправлена проблема, из-за которой не удавалось переназначение устройств, экспортированных из USB-соединений.
исправлена ошибка, из-за которой при экспорте текста иногда использовались устаревшие значения параметров
- сборка 274 [2014-02-26] -
исправлены некоторые ошибки параллелизма
- сборка 273 [2014-02-05] -
добавлена поддержка функциональных блоков Logic / 8-bit Binary Decoder и Logic / Byte Splitter.
исправлена ошибка в системе сигнализации savvyPanel.
обновлены некоторые внутренние параметры электронной торговли
- сборка 272 [2014-01-15] -
исправлена ошибка, из-за которой функциональные блоки, содержащие типы параметров «объект» (например,грамм. программ) не удалось перетащить-дублировать
исправлена ошибка, из-за которой нехватка места для программы устройства некорректно обрабатывалась для фантомов
исправлена ошибка параллелизма, которая иногда проявлялась во время импорта
добавлена поддержка программного обеспечения Optidrive P2 версии v1.30
добавлена поддержка отображения всех значений параметров в диаграммах SFD
добавлена метка времени для подписанных файлов JAR
- сборка 271 [2013-12-04] -
добавлена поддержка PL-серии версии 6.10
исправлено значение по умолчанию для параметра 79 серии PL в платах управления v2
исправлена ошибка, из-за которой маршалированный опрос не использовался во время обнаружения для dw2X1
- сборка 270 [2013-11-15] -
исправлена ошибка, из-за которой включение устройства в выбор предотвращало перетаскивание и т. д.от работы
добавлен пункт меню «Сохранить все параметры (серии PL и JL)» в меню «Каталог устройств»
добавлен пункт контекстного меню Set Graphic Location ... для элементов SFD
исправлено несколько ошибок при перетаскивании отдельных функциональных блоков
исправления и изменения для JL-серии
исправлены ошибки в "персонализированных" файлах JNLP
улучшенный вывод листинга для конечных автоматов
добавлены кнопки меню экспорта / обмена в информационное окно
- сборка 269 [2013-11-05] -
улучшен диалог "Добавить функциональный блок ..."
добавлены функции для обозначения и поддержки устаревших функциональных блоков
добавлена поддержка функционального блока Utility / Watchdog v2
добавлена поддержка PL-серии версии 6.09
добавлена поддержка ссылок между savvyPanel и устройствами в SFD
обновлен код определения модели для Optidrive E2
исправлена ошибка в редакторе профиля кулачка, при которой отображаемые координаты y были инвертированы
вставляемые примечания пользователя теперь ограничены по длине примерно до 120 символов
добавлено специальное сообщение об исключении, когда пользователь пытается подключить GOTO серии PL к GETFROM
добавлена поддержка системы присутствия savvyPanel
добавлена поддержка функционального блока savvyPanel / Presence Monitor
добавлена поддержка функции сигнализации savvyPanel
добавлена поддержка функционального блока savvyPanel / Alarm Annunciator.
добавлена ограниченная поддержка пользовательских логотипов в savvyPanel
добавлена функция «Изменить модель» для фантомов
добавлен 'Загрузить прошивку для устройств... 'в меню каталога устройств
добавлен пункт меню «Загрузить ПО серии PL ...» в меню «Каталог устройств».
добавлена функция "Изменить фантом"
настройки диалогового окна границы теперь запоминаются для фантомов
добавлена поддержка группового выбора, перемещения, перетаскивания и удаления
обновленное руководство пользователя
обновлен PL / X Quick Start
добавлена поддержка JL-серии версии 1.23
- сборка 268 [2013-08-19] -
исправлена ошибка последовательной связи для Optidrive E2
исправлено несколько проблем с руководством пользователя
исправлена ошибка для PL-серии, когда активированные GETFROM FB всегда показывались неактивными
добавлена поддержка опции -36 и функциональных блоков «Motion Control / Trapezoid Motion» и «Motion Control / Cam Profile».
добавлена поддержка опций -37 и -38 и функциональных блоков Stepper / Stepper 1, Stepper / Stepper 2 и Stepper / Stepper Status Monitor.
добавлена поддержка новой платы управления серии PL, версии 6.07 и 6.08
- сборка 267 [2013-06-27] -
исправлена ошибка в установщиках параметров для PIN 2 и 4 серии PL
добавлена поддержка Optidrive E2 версии 1.10
обновленное руководство пользователя
- сборка 266 [2013-06-19] -
обновленная система сборки и т. д. - теперь использует Java 6 (и требует Java 6 или новее)
в Mac OS X удален Quaqua filechooser (так как он больше не поддерживается)
исправлен сбой при вводе пустой строки для формата в параметре Re-Scale
добавлена поддержка функционального блока 'Ramps / Setpoint Programmer'
добавлена поддержка программного обеспечения Optidrive P2 версии 1.20
добавлена поддержка функционального блока 'Утилита / Рецепт'
добавлена поддержка функциональных блоков 'Frequency / Frequency Speed Lock' и 'Frequency / Frequency Phase Lock'
добавлена поддержка «Частота / высокочастотный вход T3», «Частотный / высокочастотный вход T4»
и функциональные блоки "Частотный / высокочастотный вход T5"
добавлена поддержка функционального блока Frequency / Frequency Edge Counter 3.
добавлена поддержка дополнительной карты цифрового входа Optidrive P2 и дополнительной карты релейного выхода
добавлена поддержка Optidrive P2 v1.20 параметров P0
исправлена ошибка, из-за которой "Очистить журнал" был доступен для фантомов (у которых нет журнала)
добавлена поддержка функциональных блоков Logic / 2-Bit Priority Encoder и Logic / 3-Bit Priority Encoder.
исправлена ошибка, из-за которой некоторые имена были слишком длинными в корзине покупок
добавлена поддержка границ ANSI C, ANSI D, ISO A2 и ISO A1.
добавлена предварительная поддержка опции -36 и функциональных блоков Motion Control / Trapezoid Motion и Motion Control / Cam Profile.
добавлена поддержка функционального блока Control / Clamped Integrator.
добавлена предварительная поддержка опций -37 и -38, а также функциональных блоков Stepper / Stepper 1 и Stepper / Stepper 2
улучшенный диалог заметок пользователя
добавлена поддержка копирования и вставки заметок в SFD
добавлена поддержка служебных / аналоговых параметров, служебных / логических параметров, служебных / перечислимых параметров.
и функциональные блоки «Служебные / шестнадцатеричные параметры»
добавлена поддержка параметров "Время или скорость"
изменен z-порядок в SFD, чтобы параметры помещались поверх соединений
добавлена поддержка преобразования формата с плавающей запятой в функциональный блок «Математика / Калькулятор»
добавлена поддержка функциональных блоков «Служебные / двоичные параметры» и «Блок служебных / двоичных параметров».
исправлена ошибка в корзине покупок, которая возникала при наличии обновлений до нескольких поколений продуктов
добавлены альтернативные блоки заголовков для некоторых размеров страниц
изменен параметр «Очистить все внешние соединения», чтобы не затрагивать соединения между FBE и устройством в локальном модуле.
- сборка 265 [2012-12-19] -
исправлены ошибки в дециматоре тренда
улучшен дециматор тренда
исправлена ошибка в коде определения модели для Optidrive E2
- сборка 264 [2012-11-16] -
исправлена ошибка, из-за которой «Импорт данных и создание фантомов» не удался, если в файле экспорта были USB-устройства
исправлена графическая ошибка для Optidrive E2, когда аналоговое масштабирование и смещение были транспонированы
добавлена поддержка функционального блока Encoder / ENC Event Counter 2
добавлена поддержка функционального блока Frequency / Frequency Edge Counter 2.
улучшил трендовый объект
добавлена поддержка PL-серии версии 5.22 и 5.23
- сборка 263 [2012-10-05] -
включена поддержка USB
добавлена поддержка Optidrive E2
улучшенная система обновления программного обеспечения
внесены незначительные улучшения в программный редактор State Machine
добавлена поддержка функционального блока «Арифметика / масштабирование»
добавлена поддержка функционального блока 'Encoder / ENC Event Counter'
добавлена поддержка функционального блока Frequency / Frequency Edge Counter.
- сборка 262 [2012-05-09] -
добавлена поддержка ODP2 версии 1.10
добавлена поддержка loopback (127.0.0.1) соединений
исправлена ошибка, из-за которой платформы на испанском, французском и немецком языках не запускались.
исправлена ошибка, из-за которой возможности фантомов dw220 (и dw210) были некорректными
исправлена ошибка, из-за которой неизвестная версия прошивки ODP2 вызывала NPE
исправлена ошибка в мониторинге асинхронных транзакций
добавлена поддержка USB (станет общедоступной в сборке 263)
исправлена ошибка в подписи кода, которая могла вызывать различные исключения "подписывающего код"
исправлена ошибка, из-за которой устройства speedy2 сообщали свой тип как "smarty2"
- сборка 261 [2012-02-23] -
добавлена предварительная поддержка накопителей серии JL
сняты ограничения по типу устройства из импорта
- сборка 260 [18.01.2012] -
добавлен индикатор выполнения в «Загрузить прошивку»
исправлена ошибка, из-за которой перетаскивание графической плитки на новую страницу превращало ее в плитку со ссылкой
исправлена ошибка, из-за которой предупреждения об обмене данными в панели savvyPanel не сбрасывались после установления связи с устройством
исправлена ошибка, из-за которой «Загрузить графику по умолчанию» не возвращал представление
внесены улучшения в пользовательский интерфейс диалоговых окон паролей и диалоговых окон перечисления пользователей.
обновлено «Создать фантомное устройство»... 'диалог
добавлена поддержка 'Library 6 Solar'
исправлены ошибки, связанные с параметрами PLX 'fieldbus'
добавлена поддержка ODP2
- сборка 259 [2011-06-29] -
исправлена ошибка в редакторе программы конечного автомата, возникавшая при удалении последнего перехода и последующем добавлении нового
добавлена поддержка функционального блока: 'Solar / Sun Position'
добавлена поддержка функционального блока: «savvyPanel / SR Latch»
добавлена поддержка функциональных блоков: «Логический / 8-канальный демультиплексор» и «Логический / 16-канальный демультиплексор»
исправлена ошибка отображения, которая проявлялась при перетаскивании плиток между окнами savvyPanel
исправлена ошибка экспорта / импорта специального типа параметра "ODP Speed"
- сборка 258 [2011-05-25] -
добавлена поддержка функциональных блоков: «savvyPanel / Monitor», «savvyPanel / Setpoint & Monitor», «savvyPanel / Dual Monitor»
добавлена поддержка функционального блока: 'Индексированная циклическая позиция кодировщика / ENC'
исправлена ошибка в пользовательском демонстрационном режиме, когда выход с возможностью SFD очищал фантомную графику
добавлена поддержка плитки savvyPanel 'Cyclic Meter'
исправлена ошибка, из-за которой единицы, введенные пользователем, не экранировались должным образом при экспорте XML
исправлена графическая ошибка в функциональном блоке 'Моталки / Компенсатор крутящего момента' субблок 'Трение'
улучшена обработка неполных записей в диалоговом окне перечисления пользователей
добавлена поддержка функциональных блоков: 'Logic / State Machine 8-8', 'Logic / State Machine 16-16'
добавлена поддержка функционального блока: «Математика / Калькулятор»
исправлена ошибка, из-за которой графические данные подключения не всегда создавались должным образом
добавлена поддержка возможности прошивки "savvyPanel"
добавлена поддержка функционального блока: 'Logic / Combinational 6-4'
- сборка 257 [2011-01-26] -
исправлена ошибка обфускации при создании фантома диска
- сборка 256 [2011-01-26] -
исправлена ошибка, при которой отображалась новая запись перечисления для полностью перечисленных параметров
исправлена ошибка, из-за которой пользовательский демонстрационный режим нельзя было отключить при определенных условиях
исправлена ошибка рисования тренда
исправлена ошибка в журнале отчетов
исправлена ошибка, из-за которой последовательные устройства не сортировались, когда был установлен параметр «Сортировать по IP»
исправлена ошибка (только для Windows), из-за которой сеттеры в savvyPanel не могли быть перемещены
добавлена поддержка функциональных блоков: «Циклическая позиция энкодера / ENC1», «Циклическая позиция энкодера / ENC2»,
«Монитор состояния кодировщика / ENC», «Индексированная позиция кодировщика / ENC», «Преобразователь циклических ошибок кодировщика / ENC»,
«Encoder / ENC Cycle Calibrator Basic», «Encoder / ENC Cycle Calibrator DIN1» и «Encoder / ENC Cycle Calibrator DIN2»
- сборка 255 [2010-11-02] -
улучшенный диалог создания фантома
поддержка обратного переноса идентификаторов групп на основе строк в продукты dw1XX
добавлена поддержка функционального блока ModbusRTU Master / Comms Monitor.
добавлена поддержка панели оператора savvyPanel
добавлена поддержка фильтрации обнаружения
добавлена поддержка пользовательских перечислений для параметров
добавлена поддержка для 'Utility / Enumerated Parameter Block' и 'Utility / Hexadecimal Parameter Block'
улучшенный диалог настроек
добавлена поддержка файла обнаружения запуска
объединил различные «Ясные»... 'через диалоговое окно
улучшенная поддержка параметров Fieldbus для серийных PL-серий
расширенная поддержка пароля устройства, позволяющая ограничить любой доступ к устройству
версия 3.5
- сборка 254 [2010-05-10] -
исправлена ошибка, из-за которой иногда отсутствовало украшение "не по умолчанию" при первом отображении параметра
исправлена ошибка, из-за которой печать многостраничных документов работала некорректно
- сборка 253 [2010-05-05] -
добавлен обходной путь для проблемы с возможностями x86 / amd64 в Windows
добавлена поддержка возможности 'EIP / PCCC Server'
исправлена ошибка регрессии при создании фантомов E-серии
исправлены мелкие ошибки в файлах JNLP
исправлена ошибка, из-за которой файл поддержки последовательной связи для 64-битной Windows был неправильно размещен
изменил способ обработки обновлений, чтобы обойти некоторые ошибки Oracle JWS.- сборка 252 [2010-03-11] -
добавлено исправление истечения срока действия SSL-сертификата сервера электронной коммерции
исправлена ошибка отображения при открытии панели «Сетевая информация» в «Настройка IP-адресов для системы»
- сборка 251 [2010-03-02] -
добавлена поддержка функционального блока 'Utility / E-Mail Notification'
исправлена ошибка в средстве перевода импорта кросс-версий для Optidrive Plus
исправлена ошибка, из-за которой фантомы dw213 с опцией -16 не могли создавать функциональные блоки кодировщика
добавлена поддержка кнопок меню при наведении
исправлена ошибка, из-за которой движения заметок пользователя не фиксировались должным образом
добавлена поддержка маршалированного опроса для dw210
улучшена производительность алгоритма построения графиков тенденций для снижения нагрузки на компьютер
улучшен диалог создания фантома ODP для разделения моделей между американскими и международными
добавлены «селекторы моделей» для фантомов Optidrive Plus и PL.
добавлена поддержка моделей серии PL 800, 900 и 980.
добавлена поддержка моделей Optidrive Plus 500-600V
добавлена поддержка установки версии прошивки при создании фантомов
- сборка 250 [17.12.2009] -
добавлена поддержка программного обеспечения Optidrive Plus версии 3.10
добавлена поддержка dw213
исправлена ошибка, при которой адрес маршрутизатора и сетевая маска были перенесены для dwM5
добавлена поддержка функционального блока Utility / Modbus Master Stats.
- сборка 249 [2009-11-19] -
дополнительно скорректирован код обнаружения модели серии PL
добавлена поддержка прошивки 0x2010
исправлена ошибка, из-за которой обновление возможностей dw210 приводило к сбою и приводило к невозможности использования устройства
- сборка 248 [2009-11-06] -
добавлена поддержка ModbusRTU Slave
добавлена поддержка функциональных блоков 'Encoder / ENC1 Position' и 'Encoder / ENC2 Position'
исправлена ошибка в шестнадцатеричных установщиках
добавлена поддержка функциональных блоков Encoder / ENC Position Point и Encoder / ENC Position Monitor.
улучшена обработка типов параметров ассоциации
добавлен «Файл-> Утилиты-> Hex Converter», чтобы упростить ручную настройку «Точки положения кодировщика / ENC».
исправлена ошибка на платформе Windows, из-за которой ассоциации не могли быть установлены
- сборка 247 [2009-10-06] -
исправлены некоторые текущие ошибки масштабирования дисплея для кадра 5 серии PL
добавлена поддержка Logic Gate / AND4, Logic Gate / OR4, Logic Gate / NAND4, Logic Gate / NOR4.
добавлена поддержка "Защелки / Т-защелка со сбросом"
добавлена поддержка ModbusRTU Master
- сборка 246 [2009-09-11] -
улучшения в поддержке "Демо-режима"
изменения в электронной коммерции для поддержки транзакций по кредитным картам с новым внутренним сервером
обновлены собственные библиотеки последовательной связи для Windows 7 и Mac OS X 10.6 Снежный барс
добавлена поддержка последовательной связи для 64-битных операционных систем
теперь используется Quaqua 5.4.1 на платформе Mac OS X (только для выбора файлов)
добавлен обходной путь для ошибки Java только для Windows, когда поля со списком не отображаются должным образом в установщиках параметров
удален временный обходной путь для "Java для Mac OS X 10.5, обновление 4"
[поскольку обе версии «Java для Mac OS X 10.5 с обновлением 5» и «Mac OS X 10.6» теперь выпущены и устраняют проблемы]
обновлена поддержка опций dw210 -16, -18, -22, -23 [i2i comms]
обновлена поддержка «I / O / ENC Speed Lock» и «I / O / ENC Phase Lock»
добавлена поддержка для 'Control / Trim' и 'Logic / Bitwise Match'
добавлена поддержка опции dw210 -24 [дополнительные цифровые входы]
добавлена поддержка 'I / O / T03 Digital Input' через 'I / O / T08 Digital Input' и 'I / O / LED Control'
улучшена автопрокрутка при перетаскивании в списках программ FBE
- сборка 245 [2009-07-16] -
добавлены отдельные «Справка-> Показать размеры функциональных блоков dw1xx» и «Справка-> Показать размеры функциональных блоков dw2xx»
добавлена поддержка dw210 опций -16, -18, -22, -23
добавлена поддержка «I / O / ENC Speed Lock» и «I / O / ENC Phase Lock»
добавлен способ решения проблемы с возможностями i386 / x86_64 в Mac OS X
исправлена ошибка, из-за которой соединения driveweb от FBE к локальному устройству отображались как неразмещенные в списке FBE
добавлен временный обходной путь для проблем, вызванных Java для Mac OS X 10.5 Обновление 4 '
ПРИМЕЧАНИЕ: на Mac меню теперь находятся в окнах - это требуется для вышеуказанного (временного) обходного пути.
- сборка 244 [2009-05-15] -
исправлена ошибка, из-за которой 'Utility / Modbus Indirect Block' был недоступен в dwM5
добавлены дополнительные предупреждения о «Демо-режиме»
добавлена поддержка OptidrivePlus версии 3.01
обновлена поддержка dw210 option -15
- сборка 243 [2009-04-22] -
добавлена поддержка опции dw210 -15
- сборка 242 [2009-03-19] -
исправлена ошибка, из-за которой после импорта иногда могла отображаться неправильная графика для функциональных блоков
исправлена ошибка, из-за которой «Клонировать в фантом» не работало.
исправлен набор ошибок на платформе Windows, из-за которых самое переднее окно могло измениться после закрытия диалога
исправлена ошибка, из-за которой лимит размера пользовательских данных устанавливался неверно для dwM5
добавлен обходной путь для ошибки в функции «Загрузить прошивку» прошивки 0x2000
возможность последовательной записи параметров GOTO fieldbus серии PL
добавлено обнаружение повторяющихся входящих подключений driveweb
добавлена проверка, чтобы "болтающиеся" исходящие соединения driveweb учитывались при создании новых соединений
добавлена поддержка OptidrivePlus v3.00; улучшена поддержка v2.20, v2.21 и v2.22
добавлена поддержка установки версии ПО устройства при создании фантомов
создание руководства пользователя теперь происходит в фоновом потоке (по мере необходимости)
исправлена ошибка, из-за которой все параметры в руководстве пользователя отображались только для чтения
теперь используется Quaqua 5.0.1 на платформе Mac OS X (только для выбора файлов)
- сборка 241 [2008-12-19] -
добавлены украшения в диалоговое окно «Экспорт», чтобы привлечь внимание к заблокированным устройствам или сбоям связи.
исправлена ошибка, из-за которой стрелка навигации "контейнер" (вверх) не работала для dw1101
обновленный сертификат безопасности на 2009 год
исправлена потенциальная ошибка взаимоблокировки в потоке связи с устройством
улучшенная отзывчивость приложения в условиях сбоя связи
включена поддержка продукта dw210 Smarty
исправлена несогласованность пользовательского интерфейса в аналоговом сеттере
обновленная система сборки для создания подписанных файлов JNLP
- сборка 240 [2008-10-10] -
изменен символ функционального блока «Защелка» серии PL, чтобы лучше показать его функциональность
добавлен обходной путь для ошибки "URL-адреса ресурсов, поврежденные в JWS" в Java 1.5.0_16 (и 1.4.2_18)
добавлено предупреждение в диалогах паролей, когда активна клавиша Caps Lock
- сборка 239 [2008-09-09] -
исправлена ошибка, из-за которой удаление внутреннего подключения накопителя серии PL не было атомарным
исправлена ошибка, при которой прототипы модели использовались по ссылке в фантомах
добавлена предварительная поддержка рам 4 и 5 серии PL
исправлена ошибка, из-за которой имена в файлах док-станции не экранировались
- сборка 238 [2008-08-07] -
добавлена настройка, позволяющая автоматически возвращаться к просмотру страницы при изменении размера окна
исправлена ошибка в 'Re-Map Dock File', когда заголовок переназначенного документа не обновлялся
исправлена ошибка, из-за которой более 32 внешних подключений FBE вызывали исключения (поддерживаются 64)
добавлено примечание к функциональному блоку приводов серии PL «Защелка», поясняющее его работу.
- сборка 237 [2008-06-20] -
добавлен лучший значок для "неизвестного типа устройства"
исправлена ошибка, из-за которой фантомные программы не всегда обновлялись сразу после изменения
исправлена ошибка, из-за которой редактирование или перемещение заметки иногда могло приводить к дублированию
добавлен новый пункт меню «Экспорт в файл изображения... '
обновлен символ сбоя связи Ethernet, чтобы соответствовать новой метке dwM5
добавлены демо-функции
изменено поведение импорта параметров для фантомов - теперь принимаются все значения
Результаты поиска параметра теперь отсортированы
добавлена поддержка функциональных блоков I / O / ENC1 Position2 и I / O / ENC2 Position2.
- сборка 236 [2008-04-03] -
исправлена ошибка, из-за которой руководство пользователя не открывалось для неанглоязычных локализаций
удален пункт меню справки для форума поддержки
добавлен экспорт CSV-файла для данных тренда
добавлена документация по функциональным блокам для всех функциональных блоков FBE в руководство пользователя
исправлена ошибка, из-за которой программы FBE на фантомных устройствах не имели временной шкалы
исправлена ошибка, из-за которой отмена удаления устройства не работала корректно
исправлена ошибка, из-за которой диалог создания фантома иногда мог неправильно позиционироваться
- сборка 235 [2008-01-30] -
исправлена пара опечаток в функциональном блоке ModbusRTU Master / Eurotherm ERCFW09
удалены все локализации с машинным переводом (которые, по-видимому, были непонятны носителям языка)
добавлена поддержка внутристраничных ссылок для контекстных страниц руководства
добавлены информационные кнопки для всех FB, у которых есть страницы руководства пользователя
добавлена поддержка параметров монитора Modbus 16-63 в привод Yaskawa F7
добавлена поддержка функционального блока ModbusRTU Master / WEG CFW09
добавлена поддержка функционального блока «Логический / 16-битный двоичный декодер»
- сборка 234 [2007-12-10] -
исправлена проблема обфускации, которая препятствовала созданию фантомов
- сборка 233 [2007-12-07] -
исправлена проблема обфускации
- сборка 232 [2007-12-07] -
масштабирование с помощью колеса прокрутки теперь центрируется на мыши, а не на окне (это можно отключить в настройках)
добавлен параметр для типа границы по умолчанию (автоматический, ISO или ANSI)
добавлен флажок "Импортировать данные устройства", чтобы разрешить импорт только пользовательских данных
добавлены более значимые сообщения для подключений, когда файл экспорта FBE импортируется в устройство, отличное от FBE.
добавлена поддержка функционального блока ModbusRTU Master / Eurotherm ERCFW09
добавлена поддержка фантома для dw121 speedy485-6pc
добавлена поддержка подкованного шлюза
исправлена ошибка синхронизации потоков, которая могла вызвать проблемы с графикой
изменен цвет текста для 'Timebase Overrun' обратно на оранжевый (E06402) с желтого (FFED2D)
- сборка 231 [2007-10-23] -
границы для графики по умолчанию теперь адаптируются к языку пользователя (ANSI A или ISO A4)
новые границы теперь по умолчанию для персонализированной версии (если есть)
- сборка 230 [2007-10-17] -
добавлена поддержка функционального блока Drive Helper / Optidrive Helper
добавлена поддержка функционального блока ModbusRTU Master / Optidrive VTC (заменяет ModbusRTU Master / Optidrive)
добавлена поддержка функционального блока ModbusRTU Master / Optidrive Plus
обновлены фото для Speedy485.
добавлена страница руководства для функциональных блоков ModbusRTU Master
перетаскивание окна теперь перемещает его на передний план
исправлена ошибка в скриптовой системе, когда некоторые строки некорректно кодировались URL-адресом
изменен цвет текста для 'Timebase Overrun' с оранжевого (E06402) на желтый (FFED2D)
фиксированное масштабирование параметра для параметра Yaskawa F7 U1-03 (недокументированное масштабирование)
- сборка 229 [2007-09-26] -
изменен код импорта для Yaskawa F7, чтобы уменьшить количество записей в EEPROM
исправлена пара проблем с базой данных параметров для Yaksawa F7
добавлена поддержка функционального блока 'Утилита / Индикатор'
добавлена поддержка трендов для большего количества типов параметров - e.грамм. Шестнадцатеричный
добавлена команда 'Re-Map Dock File'
исправлена ошибка, из-за которой «Импорт и создание фантомов» иногда не устанавливал значения некоторых параметров.
добавлена поддержка функционального блока ModbusRTU Master / Optidrive.
- сборка 228 [2007-08-31] -
исправлена ошибка синхронизации потоков, которая могла вызвать проблемы с графикой
теперь отключите меню 'File-> Open Recent', если подменю пусто
исправлена ошибка, из-за которой двойной щелчок по документу для запуска смекалки в Windows не открывал документ
добавлена поддержка функционального блока «Логический / побитовый сдвиг».
добавлена проверка максимального размера кучи JVM в код инициализации
исправлена ошибка, из-за которой измененные параметры не экспортируются / импортируются правильно
некоторые исправления базы данных параметров для Yaskawa F7
- сборка 227 [2007-08-22] -
исправлена ошибка, из-за которой в диалоге «Изменить имя устройства» использовался неправильный заголовок
в диалоговом окне «Повторно сопоставить файл экспорта» предварительно ввести текущие IP-адреса, чтобы пользователю оставалось только ввести измененные адреса
убрана проверка достижимости из процедуры создания фантома
исправлена ошибка регрессии, при которой параметры с типом данных ассоциации не могли быть доступны
- сборка 226 [2007-08-17] -
исправлена ошибка, из-за которой фантомы не могли быть созданы, если не было доступной IP-сети
- сборка 225 [17.08.2007] -
добавлен 'Настройка IP-адресов для системы... 'функция
обновленное руководство для вышеуказанного
исключена пользовательская настройка «Сетевой интерфейс для многоадресной рассылки»
добавлена техническая поддержка драйвера ModbusRTU Master
исправлена ошибка в обработке Yaskawa F7 ACCEPT / ENTER
добавлена возможность масштабирования параметров
добавлена поддержка линейки продуктов speedy485
переработан диалог "Создать фантомное устройство ..."
- сборка 224 [2007-07-27] -
убрана строгая проверка версии прошивки для Yaskwa F7
удалено 'Показать веб-страницу загрузки программного обеспечения' из меню 'Справка' (больше не требуется благодаря Java Web Start)
удалена функция проверки интернет-версии (больше не требуется благодаря Java Web Start)
добавлено автоматическое «угадывание» IP-адресов для фантомов
исправлена ошибка обработки кредитных карт для локализаций, в которых не используется символ "."десятичный разделитель
исправлена ошибка в обработке последовательных коммуникаций PLX для локализаций, в которых не используется "." десятичный разделитель
исправлена ошибка базы данных параметров в серии PL для PIN 199 и перенесен PIN 199 на зарезервированную страницу
добавлена (ограниченная) проверка на наличие повторяющихся IP-адресов
добавлена поддержка многосетевых подключений для проверки конфликтов локальных IP-адресов.
- сборка 223 [2007-07-12] -
добавлено диалоговое окно с предупреждением при попытке экспорта на устройстве при ошибке связи
исправлена ошибка в обработке персонализации
добавлено имя функционального блока на страницу Connections для параметров FBE
изменен алгоритм размещения доков (теперь слева направо, четыре ширины)
множество исправлений и улучшений предварительной поддержки Yaskawa F7
- сборка 222 [2007-05-04] -
изменил все ссылки "размер кадра" на "модель"
исправлена ошибка, из-за которой создание нескольких фантомов для ODP создавало старые фантомы Optidrive
добавлен комментарий с датой и номером сборки для экспорта файлов
обновлено оформление устройства "Ожидающие изменения"
добавлена поддержка последовательных устройств в файлах обнаружения
исправлена ошибка, из-за которой цели мыши в вариантной графике не распознавались
добавлен фильтр в диалоговое окно настроек, чтобы удалить петлевой интерфейс из доступных вариантов многоадресной рассылки
добавил несколько внутренних улучшений в DVG
добавлен пункт меню справки для отображения размеров функциональных блоков FBE
добавлена предварительная поддержка накопителей Yaskawa F7
добавлено украшение устройства для ошибок FBE
улучшенные украшения в целом и обновленная страница руководства
- сборка 221 [2007-03-28] -
исправлена ошибка, из-за которой сеттеры иногда отображались для параметров PL, которые не доступны для последовательной записи
исправлена ошибка, из-за которой при очистке журнала не очищался указатель журнала должным образом
добавлена предварительная загрузка зависимых параметров графического устройства для повышения производительности и предотвращения случаев сбоя
обновленное управление зависимостями кода опции для создания фантома
добавлен отчет о плохих графических файлах данных для последовательного обнаружения
изменен диалог «Создать фантом», чтобы исключения не закрывали диалог
улучшенная обработка пользовательских данных подключения, которые становятся недействительными из-за удалений
добавлена новая система управления ресурсами устройства
добавлена поддержка функционального блока 'Арифметика / Множитель-делитель'
добавлена графика по умолчанию для Smarty (просто граница)
фиксированный номинальный период подключения для ODP (сейчас 8 мс, было 30 мс)
исправлена ошибка в 'Ramps / S Ramp', когда параметры 'Reverse Up Time' и 'Reverse Down Time' были транспонированы
добавлен виджет для просмотра значения параметров только для чтения в SFD
добавлена легенда для графиков трендов во всплывающем меню
исправлена ошибка при сохранении дока с "фантомными" параметрами
исправлена ошибка в механизме повтора тайм-аута связи
исправлена ошибка, из-за которой период подключения для подключений из FBE отображался как запрошено, а не как фактический
добавлена поддержка функциональных блоков Utility / Device Comms Monitor и Utility / Watchdog Driver.
исправлена ошибка, из-за которой функция «Найти параметр» (и производные функции) не могла найти параметры функционального блока системы
добавлен минимальный (0.9%) «щекочущий» опрос параметра устройства, даже если список опроса содержит только параметры FBE
- сборка 220 [2007-02-07] -
добавлена поддержка функциональных блоков Utility / Parameter Block и Utility / Modbus Indirect Block.
добавлена возможность обнаруживать Smarty-odp (dw113), у которого сбой связи с диском
добавлена поддержка Windows Vista
- сборка 219 [2007-01-26] -
исправлена ошибка на странице DOP3 PL-серии
обновленный сертификат безопасности на 2007 год
исправлена ошибка, из-за которой «Импорт и создание фантомов» при последовательном экспорте добавлял возможности FBE
изменены названия корпусов для серии PL для устранения токов
улучшенный код определения кадра для серии PL
внесены изменения для поддержки "бесплатных вещей" из библиотеки FBE 5
добавлена поддержка функционального блока 'Control / PI'
улучшенное представление лицензии
обновленная лицензия
исправлена проблема, из-за которой неполное экранирование URI могло привести к сбою отчета об ошибке
работал над проблемой Windows, из-за которой длинные URI приводили к ложному сообщению об ошибке.
улучшенный код, который загружает пользовательские данные, так что количество ошибок ограничено и не сбрасываются все пользовательские данные
изменен код, чтобы разрешить отображение и редактирование программ FBE, которые внутренне несовместимы
добавлен новый ListFilter для неразрешенных подключений
добавлен новый значок списка для неразрешенных подключений
- сборка 218 [2006-12-13] -
обновлен раздел "требования" руководства и добавлены примечания об автоматических обновлениях
исправлена проблема совместимости с Java 6
обновлено предупреждение и текст лицензии
- сборка 217 [2006-10-31] -
добавлена возможность вставить параметр в SFD
исправлена ошибка в диалогах подключения PL GOTO и GETFROM, где не исключались GETFROM и GOTO
исправлена ошибка в коде определения размера кадра для Optidrive Plus v2.21 год
- сборка 216 [2006-10-24] -
добавлена информация о трассировке полного стека в отчеты об ошибках
исправлена ошибка, из-за которой некоторые диалоги не были локализованы и не могли использовать десятичный разделитель запятой
- сборка 215 [2006-10-16] -
улучшен внешний вид конечных точек для подключений к "внутренним" параметрам
добавлена страница руководства с подробным описанием ограничений последовательной связи серии PL при записи некоторых параметров
исправлена ошибка инициализации окна Coupon Manager, которая не позволяла запускать на Java 1.4
- сборка 214 [2006-10-09] -
Поддержка Optidrive Plus завершена
изменен импорт, чтобы исключения, возникающие при настройке параметров, не прерывали импорт
добавлена поддержка ваучеров в корзине покупок
добавлена справочная страница для торговли
несколько изменений для лучшей поддержки персонализации
добавлены параметры контроля тока возбуждения на страницу управления полем серии PL
- сборка 213 [2006-09-27] -
исправлена ошибка, из-за которой иногда возникало исключение IllegalStateException при удалении соединений FBE
исправлена ошибка в языковой базе данных, вызывавшая исключение NoSuchElementException при запуске в некоторых неанглийских системах
- сборка 212 [2006-09-22] -
добавлена поддержка программного обеспечения Optidrive Plus 3GV-M версии 2.21 год
исправлена ошибка при перетаскивании фб, он иногда "возвращался" в исходное место
изменен валидатор IP-адреса, чтобы обойти медленные библиотеки Windows
добавлено предпочтение для прокрутки колесика мыши (вместо масштабирования)
исправлена утечка памяти, связанная с перетаскиванием в SFD
исправлена ошибка параллелизма в процедуре рисования объектов
- сборка 211 [2006-09-15] -
исправлена ошибка, из-за которой графическая информация не импортировалась в последовательные устройства
добавлена обратная связь для импорта, когда цель не может принять все данные (например, FBE в последовательное устройство)
- сборка 210 [2006-09-01] -
множество изменений и исправлений для Optidrive Plus
- сборка 209 [22.08.2006] (только для внутреннего выпуска) -
множество изменений и исправлений для Optidrive Plus
- сборка 208 [11 августа 2006 г.] (только для внутреннего выпуска) -
добавлены полные имена параметров в графики трендов
исправлена ошибка в графиках тренда, когда значения не отображались, если тренд имел только одну точку данных
изменена последовательность запуска, так что предварительная загрузка завершается до запуска пользовательского интерфейса - это позволяет избежать условий гонки
добавлена поддержка Optidrive Plus (версия Modbus)
исправлена ошибка, из-за которой не открывались функциональные блоки Компараторы / Максимум и Компараторы / Минимум
добавлена визуальная обратная связь в установщике перечисления для значений за пределами текущего мин. / макс.
добавлена поддержка функциональных блоков Timers / One Shot и Timers / Underlap
добавлена поддержка функционального блока Filters / Moving Average Filter.
переименован функциональный блок Control / Filter в Filters / Low Pass Filter
добавлена поддержка функционального блока регистрации
добавлен выбор файла графических данных в диалоги последовательного обнаружения
улучшенная компоновка графического интерфейса для диалогов последовательного обнаружения
- сборка 207 [2006-06-23] -
добавлена поддержка опции -11, библиотека 'Encoder Control': блокировка скорости IO / ENC, позиция IO / ENC1 и позиция IO / ENC2
добавлена поддержка функциональных блоков Control / Integrator и Control / Differentiator.
добавлена поддержка функциональных блоков Comparators / Maximum и Comparators / Minimum
исправлены некоторые графические ошибки в функциональном блоке калибровки серии PL
исправлены ошибки в базе данных параметров серии PL, когда максимальное напряжение было 600В вместо 1000В
улучшенное определение кадров для приводов PLD и PLXD
добавлены примечания к мануалу по межсетевым экранам
исправлена ошибка, вызывавшая исключение при подключении к сети из SFD
исправлена ошибка, из-за которой зависимые параметры иногда отображались как NaN (знак вопроса в ромбе)
исправлена ошибка, из-за которой сеттеры на страницах SFD могли показывать старые значения
добавлена настройка для перемещения навигационных стрелок в правую часть окна (это фиксированное место под Windows)
добавлена поддержка функционального блока Clamps / Deadband
увеличено максимальное количество точек данных (изменения значений) для каждого параметра в диаграммах трендов с 3000 до 20000
улучшена эффективность некоторых внутренних алгоритмов в графике трендов
добавлена страница руководства для IO / ENC Speed Lock
исправлена ошибка, при которой при использовании масштабатора текста, отличного от 100%, области подключения в параметрах на страницах SFD не были активны
добавлен обходной путь для ограничения разрешения таймера Windows в диаграммах тенденций
небольшие улучшения в диалоге создания фантома; удалена опция фантома smarty-o
- сборка 206 [2006-04-28] -
исправлена ошибка, из-за которой флаг pendingChanges не всегда сбрасывался при открытии графического файла для последовательного устройства
добавлен код для обхода некоторых типов повреждений реестра Windows
улучшенное сообщение об исключении при исчерпании пространства пользовательских данных на устройстве
исправлена ошибка, из-за которой стрелка вверх переходила на страницу FBE вместо страницы диска для функциональных блоков PL в не-SFD-подкованных решениях
добавлено напоминание об удалении старых версий в диалог «Доступна новая версия»
улучшена обработка событий открытия файлов из ОС
добавлены ассоциации файлов для Java Web Start
фиксированное масштабирование PIN 897 серии PL для последовательных подключений
незначительные исправления в базе данных параметров серии PL
исправлена ошибка экспорта текста, при которой параметры только для чтения могли быть помечены как измененные
- сборка 205 [2006-03-27] -
исправлено выравнивание графики для функционального блока Sign & Value
исправлена ошибка, из-за которой запись резервного хранилища в DeviceFBList не обновлялась после создания нового файла резервного хранилища
обновлен диалог создания фантома, чтобы соответствовать последней схеме именования от маркетинга
- сборка 204 [2006-03-08] -
исправлена ошибка, которая проявляется при первом запуске в обходном пути, выпущенном в сборке 203
- сборка 203 [2006-03-07] -
добавлен обходной путь для проблемы на платформе Windows, где имя пользователя может изменить регистр
- сборка 202 [2006-03-03] -
исправлена ошибка в 'Re-map Export File', когда графическая информация для сетевых подключений пропадала
улучшено поведение средства обновления данных асинхронного устройства в условиях серьезных сетевых ошибок
- сборка 201 [2006-02-28] -
добавлена полная поддержка Mac OS X на Intel
- сборка 200 [2006-02-20] -
добавлено предупреждение о версии прошивки в диалоге «Обновление устройства»
исправлены ошибки маркировки клемм AOP1 и AOP2 на серии E
добавлена автоматическая загрузка JRE в.exe на платформе Windows
- сборка 199 [2006-02-09] -
скорректирован максимальный размер кучи Java до 128 МБ на основе отзывов от тестирования
добавлена поддержка функционального блока: Таймер / Осциллятор
добавлена поддержка функциональных блоков: Math / Square, Math / Cube, Math / Square Root, Math / Cube Root, Math / Reciprocal
добавлена поддержка функциональных блоков: Math / Exponential, Math / Logarithm
добавлена поддержка функциональных блоков: Math / Sine, Math / Cosine, Math / Tangent, Math / ArcSine, Math / ArcCosine, Math / ArcTangent
улучшена обработка неизвестных функциональных блоков
- сборка 198 [2006-02-02] -
исправлена ошибка, из-за которой соединения между устройством и FBE не клонировались во время «Clone To New Phantom»
исправлено диалоговое окно с предупреждением "Удалить внешние подключения"
- сборка 197 [2006-01-31] -
добавлены проверки и содержательное сообщение об ошибке при выходе из хранилища пользовательских данных
уменьшенный размер графики PL по умолчанию
увеличен максимальный размер кучи Java до 512M; обновленные требования к оборудованию в руководстве
- сборка 196 [30.01.2006] -
добавлены описания функциональных блоков для функциональных блоков серии PL
исправлена ошибка генерации хеш-ключа для соединений, из-за которой графика соединения могла "пропадать"
исправлена ошибка, при которой значения параметров FBE не клонировались
добавлена поддержка оповещения о сбое связи на странице SFD
добавлены проверки на коллизии для вводимых вручную адресов FB
- сборка 195 [17.01.2006] -
добавлена поддержка программного обеспечения серии E версии 4.7
- сборка 194 [2006-01-13] -
исправлена ошибка в коде, который восстанавливается после поврежденных ключей возможностей
Улучшения пользовательского интерфейса диалогового окна создания фантома
изменен порядок сортировки функциональных блоков серии PL в SFD для соответствия ручному
добавлено больше фильтров для функциональных блоков серии PL в SFD, чтобы соответствовать меню HMI привода
- сборка 193 [2006-01-06] -
исправлена ошибка, из-за которой диалог создания фантома некорректно отображался под Windows
- сборка 192 [05.01.2006] -
различные мелкие изменения в графике PL
добавлена возможность предварительного просмотра перед печатью
исправлена ошибка, из-за которой графики трендов некорректно печатались под Windows
добавлена поддержка редактирования программы DnD FBE
включен онлайн-процессинг кредитной карты
улучшен диалог создания фантома и привязан к каталожным номерам
версия 3.4
- сборка 191 [2005-12-14] (только для внутреннего выпуска) -
объединенные диалоговые окна масштабирования и сетки в диалоговое окно «Установить границу ...»
в Mac OS X, теперь используется Quaqua 3.4.1 (только для выбора файлов)
добавлена фильтрация списков в ЮФО
исправлена ошибка, из-за которой при использовании кнопки возврата для SFD терялось состояние масштабирования для страниц, отличных от страницы 1
добавлена клавиша-акселератор 'R' в пункты меню Trend Run / Hold
добавлен "масштабирование по центру курсора"
полная поддержка PL SFD
добавлены списковые фильтры в ЮФО
добавлены отдельные шкалы меток fb и разделения соединений для данных документа
версия 3.4b1
- сборка 190 [16.11.2005] (только для внутреннего выпуска) -
исправлена ошибка, при которой данные диска PLX 5/12 экспортировались без информации о кадре
исправлена ошибка, из-за которой фокус клавиатуры отдавался фрейму (а не содержащейся графической панели) при закрытии сеттера
улучшенные строки наведения и информационные строки для подключений
исправлена ошибка, из-за которой не-PL параметры не фильтровались из списков подключений для параметров PL GOTO и GETFROM
улучшена обратная связь при перетаскивании при подключении параметров PL GOTO и GETFROM в SFD
исправлена ошибка, из-за которой сеттеры некорректно размещались в SFD
добавлена поддержка установщика для функциональных блоков в SFD
добавлена поддержка динамической графики в SFD
изменены значки и местоположения автозапуска на основе отзывов пользователей
исправлена ошибка, из-за которой исходная граница не импортировалась / экспортируется правильно
добавлена поддержка "медленного" (сдвиг) и "быстрого" (альтернативного) панорамирования клавиатуры.
улучшено поведение автопрокрутки при первом перетаскивании в окно
исправлена ошибка в расчете приращения прокрутки блока списка программ
добавлена поддержка синтетических данных (в первую очередь для использования в вариантной графике)
добавлена поддержка ссылок на руководство пользователя в DVG
добавлена поддержка фиксированных подключений
улучшено восстановление ошибок тайм-аута связи для последовательных устройств
добавлено управление состоянием полос прокрутки списка программ на графических страницах
добавлена частичная поддержка PL SFD
- сборка 189 [2005-10-17] -
добавлена функция отчета об отладке
удалено сохранение / вызов расширенного состояния окна из InfoWindow
обратите внимание, что в этой сборке отключена электронная торговля через Интернет (кредитная карта); оффлайн электронная коммерция доступна
- сборка 188 [2005-10-13] (только для внутреннего выпуска) -
изменили порядок некоторых меню, чтобы лучше поддерживать электронную коммерцию
исправлена ошибка, из-за которой диалоговое окно с первоначальным паролем нельзя было использовать для повторных попыток
исправлена ошибка, из-за которой заметки пользователя нельзя было перетаскивать между страницами
исправлена ошибка в коде обновления FBE10xx
исправлена ошибка, из-за которой '"' не экранировался в URL-адресе отчета об ошибке
исправлена ошибка, из-за которой WinME обрабатывался как WinNT w.r.t. его командная оболочка
добавлена поддержка электронной коммерции, включая корзину покупок и менеджер купонов
исправлена ошибка скейлера для параметра 897 серии PL
добавлена базовая поддержка импорта и создания фантомов для файлов экспорта с адресами последовательного порта.
добавлена поддержка функциональных блоков прошивки 0x1021:
отслеживание и удержание, 4-битный кодер с приоритетом, 4-битный двоичный кодер, 16-битный двоичный кодер,
побитовое И, побитовое ИЛИ, побитовое исключающее ИЛИ, побитовое НЕ
изменено форматирование шестнадцатеричного типа, чтобы всегда использовать верхний регистр
исправлена ошибка, из-за которой старые файлы экспорта (без данных кадра) импортировались в фантомы со значком «неизвестно»
работал над сомнительными метриками шрифта для некоторых малоизвестных символов (например,грамм. "пуля")
добавлена поддержка и предпочтение для настройки сетевого интерфейса, используемого для многоадресной рассылки
- сборка 187 [2005-09-28] -
исправлена ошибка, из-за которой диалоговое окно с уведомлением об обновлении могло конфликтовать с модальными диалоговыми окнами под Windows
изменена политика импорта / экспорта, теперь неактивные параметры FBE игнорируются
- сборка 186 [2005-09-28] -
переписаны алгоритмы диаграммы трендов для повышения производительности
изменена активация информации о точке тренда с щелчка мыши на контекстное меню
добавлена лицензия и предупреждение в диалоге начального пароля для костюмов
улучшенный алгоритм добавления подключений к программам FBE
улучшенная обратная связь пользовательского интерфейса во время перетаскивания страницы на страницу в SFD
обновлены значки для приводов серии PL и приводов серии E
добавлен отчет об исключениях для отсутствующего Serialio.файл jar
исправлена ошибка, из-за которой количество страниц иногда не обновлялось в SFD
- сборка 185 [2005-09-13] -
исправлена ошибка, из-за которой строки не экранировались должным образом в файлах экспорта XML
- сборка 184 [2005-09-13] -
внесены изменения в калькулятор ускорения для мотальных машин / компенсатора крутящего момента [за TLC]
несколько исправлений ошибок для схемы потока сигналов
- сборка 183 [2005-08-31] -
исправлена ошибка, из-за которой вариантная графика могла затмевать параметры динамического поведения
исправлена ошибка синхронизации потоков, которая иногда могла мешать импорту работать с FBE
исправлена ошибка, из-за которой окно каталога не сортировалось должным образом при обнаружении
исправлена ошибка, из-за которой курсоры перетаскивания не работали под Windows
исправлена ошибка, из-за которой записи нельзя было перетащить, чтобы перестроить док
исправлена ошибка, при которой графики трендов не обновлялись
исправлена ошибка, из-за которой преобразование внутреннего имени файла в URL-адрес не экранировалось должным образом
добавлена сетка привязки для доков
полная переработка средства "Get Info"
исправлена ошибка, при которой скрытые параметры включались в документы "экспорт в текст"
исправлена ошибка, из-за которой параметры Modbus включались в документы «экспорт в текст», когда на устройстве не было Modbus
улучшен импорт для последовательных устройств
исправлена ошибка, из-за которой имена страниц могли не отображаться в пунктах меню "перейти к"
первая сборка версии 3.3
полная поддержка схемы потока сигналов для движка функциональных блоков
- сборка 182 [2005-08-09] -
исправлено несколько ошибок базы данных параметров серии PL
исправлена ошибка присоединения к потоку, из-за которой параметры FBE оставались по умолчанию во время импорта в фантомы
- сборка 181 [2005-07-08] -
исправлены некоторые ошибки базы данных параметров серии PL, появившиеся в сборке 180
- сборка 180 [2005-07-06] -
исправлена ошибка базы данных параметров серии PL на PIN 20
добавлена поддержка параметров для полевых шин в серии PL
начальная поддержка схемы прохождения сигнала
несколько внутренних изменений в движке рисования
несколько внутренних изменений в системе событий
добавлена поддержка PL-серии v5.15 прошивка
- сборка 179 [2005-06-06] -
внутренний выпуск
исправлен ввод / вывод DVG для использования имени пользователя вместо статического имени
- сборка 178 [2005-06-03] -
исправлена ошибка в 'Re-Map Export File', из-за которой соединения $ local вызывали исключение нулевого указателя
исправлена ошибка, из-за которой автоматическая установка часов не работала
добавлен бэкэнд "средства пользовательских данных"
добавлена базовая возможность переименования параметров и функциональных блоков
добавлен экран-заставка
- сборка 177 [2005-05-25] -
исправлена ошибка, из-за которой при импорте в PL не удавалось установить значения параметров, для которых были существующие внутренние соединения
- сборка 176 [2005-05-23] -
добавлена подсистема экспорта в текстовый файл
добавлен "щекочущий" опрос для обнаружения сбоев связи с устройством при пустом списке опроса
добавлен специальный фильтр в индикатор сбоя связи устройства OptiDrive для обработки синтетических параметров
исправлена ошибка, при которой параметры в доке не удалялись при удалении содержащего их fb
добавлена команда 'Re-Map Export File'
добавлена основная поддержка электронной коммерции
изменен параметр 40151 серии E (Текущая программа Vysta) на доступ только для чтения
- сборка 175 [2005-04-26] -
фиксированные обозначения терминала кодировщика
обновленное руководство, чтобы использовать разумное имя
обновлены имена исполняемых файлов, чтобы использовать смекалку
- сборка 174 [2005-04-25] -
исправлена ошибка, из-за которой перетаскивание параметра в доке могло оставлять след на экране
добавлена поддержка прямой последовательной связи для серий PL и E
улучшенная поддержка OptiDrive
изменен алгоритм асинхронного опроса для учета времени отклика (в первую очередь для OptiDrive и последовательного порта)
использование нового упаковщика приложений для Windows
исправлены некоторые ошибки в функциональном блоке Winder / Torque Compensator
добавлена проверка, чтобы параметры FBE пропускались при импорте, если в устройстве нет FBE
исправлена ошибка, из-за которой входящее соединение не удалялось при сбое создания соответствующего исходящего соединения
теперь используется Quaqua версии 2.82 в Mac OS X
добавлена базовая поддержка фантомов OptiDrive
улучшено сообщение об ошибке, если пользователь оставляет поле IP-адреса пустым при создании фантома
добавлена поддержка локального символа для соединений в файлах экспорта
добавлен код для автоматической настройки периода фонового опроса для адаптации к условиям сети
исправлены некоторые проблемы синхронизации потоков, которые вызывали временные задержки при открытии DVG
- сборка 173 [2005-03-22] -
добавлена поддержка функциональных блоков Ramps / S Ramp и Ramps / MOP
изменен листинг fbe для использования символических имен fb
несколько мелких исправлений в графике серии E и базе данных параметров
изменено имя Clamp / Deadband fb на Clamp / Skipband
на платформе Mac OS X включен показывать Growbox
- сборка 172 [2005-03-17] -
исправлена ошибка в процедуре рисования строки, которая могла вызвать исключение нулевого указателя
включает производственный калибровочный код для Smarty
- сборка 171 [2005-03-15] -
в базе данных параметров серии E, включены разрешения на подключение для I7c - I7f
- сборка 170 [2005-03-14] -
исправлена ошибка синхронизации потоков при импорте программы и параметров FBE
исправлена ошибка в "Очистить программу", когда связанные соединения не очищались
исправлена ошибка, из-за которой потоки поллера не всегда завершались при удалении устройства
исправлена ошибка отрисовки экрана, из-за которой при динамическом обновлении строк экран оставался непослушным
улучшенные данные Get Info для внутренних соединений серии PL
улучшенный диалог исключения
на платформе Mac OS X принудительная установка расширений файлов во время сохранения (как в Windows)
различные исправления функционального блока Winder
обновлены изображения Smarty и Optidrive
добавлено имя функционального блока к параметрам 'док-станции', когда это применимо
исправлена ошибка, из-за которой адреса для новых фантомов не проверялись
изменен алгоритм сортировки для E-серии, так что параметры FBE сортируются в id-порядке
- сборка 169 [2005-02-23] -
изменено обозначение терминала Smarty, чтобы оно соответствовало производственным единицам
добавлена поддержка функционального блока Utility / Watchdog
добавлена поддержка параметров "заполнитель"
- сборка 168 не выпущена -
- сборка 167 [2005-02-01] -
исправлены некоторые проблемы с производственной тестовой лабораторией
добавлена поддержка функциональных блоков кодировщика FBE
добавлен пункт меню "Запросить улучшение"
- сборка 166 [2005-01-26] -
обновленный сертификат безопасности на 2005 год
на платформе Mac OS X, теперь с использованием внешнего вида Quaqua
после первого запуска для возможности теперь установлено значение «Администрирование», а не «Просмотр».
исправлена ошибка, из-за которой экземпляры одноэлементных FB могли быть созданы через диалоговое окно «Добавить функциональный блок».
добавлена возможность обновления возможностей функционального блока с помощью кода
добавлено примечание к руководству о внешних подключениях от GOTO серии PL и к GETFROM
добавлена производственная испытательная установка
исправлена ошибка NPE, возникавшая при открытии сохраненной док-станции, содержащей неоткрытые устройства
добавлена возможность отладки параметров
обобщенные одноэлементные проверки FBE с помощью флагов ресурсов
- сборка 165 [2005-01-05] -
исправлена ошибка экспорта возможностей
исправлена ошибка импорта данных FBE через 'Импорт и создание фантомов... '
добавлена поддержка дополнительных функциональных блоков FBE
исправлен экспорт / импорт программы FBE для использования адресации пользователей
добавлена базовая поддержка параметров ссылки функционального блока FBE
- сборка 164 [2004-12-09] -
добавлен пункт «Импорт и создание фантомов ...» в меню «Каталог»
добавлено украшение PL-серии для "ожидающих изменений параметров"
добавлена проверка IP-адреса в «Обнаружение устройства ...»
добавлен пункт меню «Очистить все журналы устройств» в меню «Каталог»
сделал пункт меню «Очистить журнал» общедоступным в контекстном меню устройства.
добавлена поддержка FBE в Clone to New Phantom..." элемент
переработана обработка FBE функциональных блоков ввода / вывода
- сборка 163 [2004-11-16] -
исправлена ошибка блокировки потока, которая могла возникнуть на странице списка подключений
- сборка 162 [2004-11-16] -
добавлена поддержка программного обеспечения серии PL версии 5.14
различные дополнения и исправления ошибок в FBE
- сборка 161 [2004-11-04] -
исправлена ошибка, при которой не отображались пункты назначения GOTO серии PL
фиксированные строки перечисления для PIN 704 серии PL
фиксированное отображение битового поля на странице Run Digital Input для серии PL
исправлено размещение параметров на страницах "Компаратор 1" - "Компаратор 4" серии PL
исправлена ошибка базы данных параметров на PIN 476 PL-серии
изменения в (обобщении) схемы генерации символа функционального блока FBE
добавлена поддержка дополнительных функциональных блоков FBE
перешли на даты ISO 8601 для этого документа
- сборка 160 - 29окт04 -
исправлена проблема обфускации, из-за которой фантомы не работали
- сборка 159 - 29окт04 -
первая сборка версии 3.2
добавлен движок функциональных блоков (с прошивкой 0x10XX)
полная переработка системы управления соединениями
значительный рефакторинг системы управления параметрами
исправлены ошибки в команде dwM1 'Получить подробную информацию'
глобально измененная эвристика периода подключения
- сборка 158 - 30ауг04 -
внутренние соединения на PL теперь используют перемычки, если они доступны
исправлена ошибка, из-за которой в PL GETFROM не отображалось "модифицированное" украшение
исправлена ошибка перетаскивания, связанная с Mac OS X 10.3 Java 1.4.2 Update 1
добавлена проверка на пустой текст в аналоговом сеттере
исправлена ошибка, из-за которой вариантная графика вызывала дополнительные ссылки для подключения
базовая поддержка автоматической генерации программы FBE (внутренние соединения)
- сборка 157 - 9ауг04 -
добавлена проверка блокировки для кода автонастройки времени
обновил различные вещи, чтобы использовать "умное" название
обновлено множество страниц PL-серии
- сборка 156 - 16 июн04 -
поддержка альфа-уровня для регистрации устройства прошивки 0x9X
добавлена поддержка динамических диаграмм
добавлена возможность сбросить информацию об IP-адресе до заводских настроек по умолчанию
обновлено диалоговое окно "Установить IP-адрес".
- сборка 155 - 19 мая 04 -
добавлены дополнительные проверки корректности IP-адреса маршрутизатора и маски подсети
улучшенные пользовательские сообщения об ошибках устройства
поддержка альфа-уровня для часов реального времени прошивки 0x9X
- сборка 154 - 12 мая 04 -
переработано украшение выхода из строя связи; добавлено отдельное украшение для внутренней связи устройства
добавлена поддержка DVG для условных графических сущностей
добавлена поддержка DVG для символьных атрибутов контейнера
исправлена ошибка масштабирования параметра в PIN 146 серии PL
«Найти параметр» теперь также выполняет поиск меток параметров (в дополнение к идентификаторам и именам)
поддержка альфа-уровня для прошивки 0x9X, включая ModbusTCP
- сборка 153 - 14апр04 -
исправлена ошибка, из-за которой фокус клавиатуры терялся при открытии аналогового сеттера
а затем закрыли без установки значения
исправлены ошибки в коде обработки кадров серии PL
предварительная поддержка Optidrive
- сборка 152 - 13апр04 -
исправлена ошибка, из-за которой установка IP-адресов могла вызвать исключение нулевого указателя
- сборка 151 - 7апр04 -
исправлена ошибка, из-за которой импорт мог завершиться неудачно для серии E при попытке зафиксировать переходные параметры
- сборка 150 - 6апр04 -
предварительная поддержка dwM4-S
добавлена маска подсети и IP-адрес маршрутизатора в устройство Get Info
добавлено обратное значение для скейлера в аналоговый параметр Get Info
переписал проверку лимита количества подключений для обработки всех ограничений
- сборка 149 - 16мар04 -
исправлена ошибка, при которой «Сортировать по MAC-адресу» и «Сортировать по номеру группы» в каталоге устройств.
окно во время обнаружения могло вызвать повреждение некоторых структур данных приложения
- сборка 148 - 15мар04 -
добавлена проверка диапазона в 'Установить группу... '(используется для усечения значений, выходящих за пределы допустимого диапазона)
- сборка 147 - 5mar04 -
добавлена поддержка E-серии версий 4.2, 4.3 и 4.4
улучшена обработка номеров версий программного обеспечения фантомных устройств
под Windows, теперь с использованием "системного внешнего вида"
- сборка 146 - 7feb04 -
в серии PL исправлена ошибка параметра, когда PIN 64 был доступен для записи
в PL-серии обновлена страница Speed Control и UIP3; добавлена страница адаптации Speed Loop PI
- сборка 145 - 13январь 04 -
добавлен пункт «Показать веб-страницу загрузки программного обеспечения» в меню «Справка»
обновленный сертификат безопасности на 2004 год
добавлены примечания к руководству о многосетевых компьютерах и локальной многоадресной рассылке
добавлена дата сборки в диалог "о"
- сборка 144 не выпущена -
- сборка 143 - 09 декабря 03 -
исправлена ошибка исключения нулевого указателя при открытии файлов док-станции, содержащих неоткрытые устройства
исправлена ошибка, из-за которой поддельное имя устройства могло отображаться в заголовке фрейма каталога устройства
обновлена страница компенсатора крутящего момента серии PL
обновлены страницы UIP серии PL
обновлены страницы DOP серии PL
обновлены страницы переключателей серии PL
обновленные многофункциональные страницы серии PL
изменено диалоговое окно "Установить соединение", чтобы использовать указанный порядок сортировки устройств.
добавлена метка параметра (если есть) в строку наведения
установить начальный фокус на поле ввода текста в аналоговом сеттере
добавлена настройка, позволяющая отключить проверку версии при запуске
исправлена ошибка, из-за которой вновь созданный и сохраненный док не мог быть просмотрен через историю страницы
- сборка 142 - 02dec03 -
исправлена ошибка, из-за которой приложение зависало при заполнении буфера тренда
исправлена ошибка гонки в коде проверки конфликта соединений серии PL
- сборка 141 - 20нов03 -
изменен диалог обновления при запуске из JWS, чтобы избежать путаницы
добавлена поддержка фоновых обновлений DObjects при обнаружении устройств
добавлена строка "содержащее устройство" в параметр "Get Info"
- сборка 140 - 17нов03 -
исправлена ошибка масштабирования в базе параметров серии PL для PIN 146
исправлена ошибка, из-за которой зависимые параметры иногда отображали неверные значения
исправлена ошибка в аналоговом сеттере зависимых параметров
исправлено несколько ошибок, связанных с фреймами, в базе данных параметров серии E
изменено форматирование в базе данных параметров серии E, поэтому десятичные цифры отображаются всегда
- сборка 139 - 13нов03 -
улучшенное поведение format (), когда данные еще не доступны
добавлена поддержка заводской автоматической сериализации
обновлен раздел требований руководства
скорректирован порядок строки меню, чтобы лучше соответствовать HIG
исправлена ошибка на странице нулевых блокировок серии PL
незначительное обновление базы данных параметров серии PL
исправлена ошибка в коде масштабирования бегущей строки, когда использовалось целочисленное деление вместо числа с плавающей запятой
обновлена графика нескольких страниц PL-серии
изменено поведение клавиш со стрелками и обновлено руководство
- сборка 138 - 02нов03 -
исправлены ошибки на страницах AOP3 и AOP4 серии PL
- сборка 137--
исправлена ошибка (регресс) в коде проверки соединения
- сборка 136--
изменены структуры данных информации о параметрах для обработки типов данных float и double в дополнение к INT16
переработана обработка подписанных INT16 и логических значений
в нескольких местах заменены перечисления в стиле c на типы безопасного типа
исправлена ошибка в битовых полях PL-серии, когда имена битов могли обрабатываться как перечисления
- сборка 135--
изменена обработка логического типа для включения PL - любое ненулевое значение теперь обрабатывается как истинное
- сборка 134--
добавлена поддержка сценариев для 'set-address'
- сборка 133--
исправлена ошибка гонки в потоке предварительной загрузки данных внутреннего соединения серии PL
- сборка 132--
добавлена функция, так что если последнее закрытое окно развернуто, приложение запускается с начальным
развернутое окно (на платформах, поддерживающих явно развернутые окна, например.грамм. Windows)
- сборка 131--
добавлен отдельный пункт меню «Параметры страницы ...» и сделаны постоянные параметры настройки страницы
--ПРЕДУПРЕЖДЕНИЕ--
из-за проблем с базой данных параметров серии PL в сборке 129 и ранее,
любые системные файлы, сохраненные от фантомов, созданных с помощью сборки 129 или более ранней, должны
быть выброшенным и воссозданным (или отправленным в Бардац для ремонта). Некоторые скрытые
параметры были установлены на значения, которые препятствуют работе текущего цикла.
(Сборка 130 исправила проблемы и была выпущена 2 октября 2003 г.)
- сборка 130--
доработано поведение "не по умолчанию", теперь параметры входящих подключений не украшаются.
добавлена поддержка продукта PLA
добавлено примечание к инструкции относительно импорта в PL-серию PIN-кодов 2 и 4
исправлена еще пара проблем с базой данных параметров серии PL
добавлено примечание к инструкции по внутренним соединениям серии PL
добавлена поддержка отображения параметров битового поля серии PL (как 8-битных, так и 16-битных)
изменена страница «Калибровка» серии PL, чтобы включить некоторые дополнительные параметры
исправлена графическая ошибка, из-за которой некорректно рассчитывалась область обновления для строк
- сборка 129--
исправлено несколько мелких опечаток в руководстве
исправлена ошибка, из-за которой параметры в фантомах долго обновлялись
- сборка 128--
добавлен мьютекс в код, который строит структуру данных документа DVG для обработки динамических изменений
исправлена еще пара проблем с базой данных параметров серии PL
- сборка 127--
добавлено примечание к ручному предупреждению об импорте на другой IP-адрес
добавлено примечание к руководству, предлагающее использовать ДЖАМПЕР, а не привод.веб-соединения в серии PL
исправлено несколько ошибок в базе данных параметров серии PL
переработан код размера кадра; добавлено меню выбора размера кадра для фантомов
исправлена ошибка, из-за которой, если параметр имел как исходящее соединение driveweb, так и внутреннее исходящее соединение
это будет отображаться как дубликат
добавлено украшение для записываемых параметров, указывающее "не по умолчанию"
обновленное руководство для вышеуказанного
- сборка 126--
исправлена ошибка в базе данных параметров серии PL для PIN 899 - неправильное значение по умолчанию и диапазон
добавлен индекс назначения для исходящих соединений в 'Получить подробную информацию'
исправлена ошибка в PL-серии, при которой непрочитанная информация GOTO отображалась как повторяющиеся соединения во время импорта
исправлена ошибка многопоточности при опросе устройства (определение смены имени / пароля / группы / соединения)
- сборка 125--
добавлен код для обеспечения видимости параметров в списке подключений при использовании навигации с помощью клавиатуры
изменено обнаружение DTD, чтобы разрешить пустой элемент верхнего уровня
убрано ограничение из "Создать файл обнаружения", где были исключены фантомы
список устройств в 'Импортировать данные... 'диалоговое окно теперь сортируется с использованием того же компаратора, что и каталог устройств.
список устройств в диалоговом окне «Экспорт данных ...» теперь сортируется с использованием того же компаратора, что и каталог устройств.
данные файла импорта теперь сопоставляются с устройствами только по IP-адресу (а не по имени и IP-адресу)
добавлены номера клемм на страницы «Инженерная информация» как для серии PL, так и для серии E
исправлена ошибка на странице AOP3 серии PL - неправильный физический номер терминала
- сборка 124--
исправлена ошибка, из-за которой не отображалось подключение параметров источника для внутренних GETFROM серии PL
- сборка 123--
обходной путь для ошибки в Mac OS X с эффектом "матового металла"
- сборка 122--
исправил пару мелких ошибок в мануале
исправлена ошибка масштабирования для параметра 489 в серии PL
исправлена ошибка в PL-серии, где параметры 691 и 692 и 693 и 694 были перенесены
обновлена поддержка Mac OS X Java 1.4.1 Обновление 1
- сборка 121--
Версия 3.1
32 предустановки памяти
Используя программное обеспечение DSP Configurator, доступно до 32 предустановок памяти для хранения и вызова настроек обработки DSP, уровней звука и микрофона, а также связей ввода / вывода аудио и видео. Также могут быть созданы предустановки только для аудио / видео связей.
Интуитивно понятная графическая среда пользователя
Программное обеспечение DSP Configurator имеет графическую среду пользователя, которая предлагает четкое представление всех входов и выходов, блоков обработки звука, связей маршрутизации аудио и видео, а также точек микширования микрофонов в одном окне.Это позволяет разработчику или установщику быстро просматривать все действия, связанные со звуком и видео, без доступа к подокнам или подменю.
Связи ввода / вывода
Графическая среда пользователя программного обеспечения DSP Configurator обеспечивает четкое представление обо всех связях ввода / вывода аудио и видео. с помощью мыши или клавиатуры связи для аудио- или видеосигналов или и того, и другого могут быть созданы, изменены или удалены в реальном времени. Затем они могут быть сохранены и вызваны как предустановки в программном обеспечении DSP Configurator.Предустановки также можно вызывать с передней панели MPX Plus 866 A, через последовательный порт RS-232 или через IP Link Ethernet.
Dynamics
Программное обеспечение DSP Configurator обеспечивает точную настройку и регулировку динамики всех входящих и исходящих сигналов от MPX Plus 866 A. Два блока обработки динамики доступны для каждого входа и выхода. Эти блоки можно выбрать и настроить, чтобы обеспечить автоматическое сжатие с регулировкой усиления, ограничение или шумоподавление.
Входные и выходные фильтры
Блок фильтров предлагает три настраиваемых фильтра для каждого линейного входа, пять для каждого микрофонного входа и девять для каждого из шести выходов. Фильтр можно выбрать из параметров параметрического эквалайзера, низких частот, высоких частот или полочного регулирования низких и высоких частот. Настраиваемые параметры включают частоту, крутизну спада, усиление / ослабление и добротность, в зависимости от конкретного фильтра.
Подавление обратной связи
Блок подавления обратной связи FBS используется для противодействия звону из-за того, что частоты выходят из-под контроля через микрофон и динамики.Процессор подавления обратной связи для MPX Plus 866A задействует до двадцати режекторных фильтров с регулируемой добротностью. Пятнадцать фильтров и динамических фильтров, и процессор использует их для автоматического обнаружения, а затем устранения звона. Пять дополнительных фиксированных фильтров можно настроить вручную или перенести из динамических фильтров.
Emulate и Live предлагают гибкие возможности конфигурации системы независимо от того, подключена ли она к коммутатору или нет.
Output Volume
MPX Plus 866A обеспечивает регулировку выходной громкости, а также отключение звука для каждого из шести A / V выходов, а также измерение в реальном времени.Для контроля любого или всех уровней входного и выходного сигналов также доступно окно измерительного моста.
Delay
Блок обработки задержки доступен для каждого входа и выхода. Задержка регулируется до 200 мс и может быть выбрана в единицах времени, футах или метрах. Параметр температуры доступен для регулировки расстояния.
Микрофонный / линейный матричный микшер
Программное обеспечение DSP Configurator также используется для управления и контроля матричного микширования микрофонного / линейного звука.Каждый микрофонный / линейный вход можно микшировать с каждым из шести доступных линейных выходов с выделенным уровнем стереомикса и элементами управления приглушением на каждом узле.
Альтернативные представления ввода-вывода
Одним нажатием функциональной клавиши пользователь может переключаться между видами ввода-вывода только аудио, только видео или аудио и видео. Программное обеспечение DSP Configurator позволяет в реальном времени изменять связи маршрутизации аудио и видео с помощью нажатия клавиш или мыши. В представлении ввода-вывода только видео доступны параметры, относящиеся к видео, включая перекодирование композитного и S-видео, задержку вывода RGB и отключение видеовыхода.
Ducking
Блок Ducking используется для присвоения приоритета определенному микрофону путем автоматического ослабления уровней других микрофонов и программных источников всякий раз, когда активен приоритетный микрофон. Настройка приглушения выполняется для всех микрофонных / линейных входов на уникальной странице глобальной настройки. Доступен широкий спектр настраиваемых опций для ныряния.
Нарушение взаимодействия LILRB4 / APOE с помощью эффективного гуманизированного антитела отменяет подавление Т-клеток и блокирует развитие ОМЛ
Abstract
Пациентам с острым миелоидным лейкозом (ОМЛ) срочно необходимы терапевтические стратегии.Лейкоцитарный иммуноглобулиноподобный рецептор B4 (LILRB4), который подавляет активацию Т-клеток и поддерживает тканевую инфильтрацию клеток AML, представляет собой привлекательную лекарственную мишень для терапевтических средств против AML. Здесь мы сообщаем об идентификации и разработке LILRB4-специфических гуманизированных mAb, которые блокируют активацию LILRB4. Это mAb, h228-3, показало сильную активность в блокировании развития моноцитарного AML на различных моделях, включая ксенотрансплантат мышей, полученных от пациентов, и сингенных иммунокомпетентных мышей AML.MAb h228-3 усиливали эффективность химиотерапевтического лечения против AML, стимулируя мобилизацию лейкозных клеток. Механистические исследования выявили четыре согласованных способа действия анти-AML-активности h228-3: (i) обращение супрессии Т-клеток, (ii) ингибирование инфильтрации моноцитарных клеток AML в ткани, (iii) антителозависимая клеточная цитотоксичность и (iv) антителозависимый клеточный фагоцитоз. Следовательно, нацеливание антитела на LILRB4 представляет собой эффективную терапевтическую стратегию для лечения моноцитарного AML.
Введение
Острый миелоидный лейкоз (ОМЛ), наиболее распространенный острый лейкоз у взрослых, характеризуется клональной пролиферацией незрелых миелоидных гемопоэтических клеток в костном мозге, крови и других тканях (1). Каждый год в США появляется 19 000 новых случаев AML, и около 10 000 случаев смерти связаны с AML (2). Несмотря на более глубокое понимание биологии, лежащей в основе ОМЛ, стандартное вмешательство цитотоксической химиотерапии не изменилось за последние 40 лет.70% пациентов в возрасте 65 лет и старше умирают от болезни в течение года после постановки диагноза (3). Более того, иммунотерапия, такая как стратегии нацеливания на CTLA4 и PD-1 / PD-L1, не принесла клинических преимуществ у пациентов с AML (4). В 2017 и 2018 годах FDA одобрило несколько новых терапевтических средств против AML, включая ингибиторы IDh2, IDh3 и Flt3, инкапсулированные в липосомы химиотерапевтические препараты и конъюгаты анти-CD33-лекарства, которые могут принести пользу некоторым подгруппам пациентов с AML (5-7). . Тем не менее, сохраняется острая необходимость в разработке новых методов лечения с высокой терапевтической эффективностью и низкой токсичностью для различных подтипов ОМЛ.
Лейкоцитарный Ig-подобный рецептор подсемейства B (LILRB) представляет собой группу трансмембранных гликопротеинов типа I, характеризующихся внеклеточными Ig-подобными доменами для связывания лиганда и внутриклеточными иммунорецепторными ингибиторами на основе тирозина (ITIM), которые могут рекрутировать тирозинфосфатазы SHP- 1, SHP-2 или инозитолфосфатаза SHIP (8, 9). Из-за их иммунно-ингибирующих функций LILRB считаются белками иммунных контрольных точек (8). Фактически, LILRB действуют на более широкий спектр типов иммунных клеток, чем классические белки иммунных контрольных точек CTLA-4 и PD-1 (10).Мы идентифицировали LILRB2 как рецептор гормона Angptl2 (11). Затем мы продемонстрировали, что дефицит мышиного ортолога LILRB2, PirB, в моделях AML приводит к усилению дифференцировки и снижению самообновления стволовых клеток лейкемии (11). Кроме того, мы и другие продемонстрировали, что несколько LILRB и родственный ITIM рецептор LAIR1 поддерживают развитие AML (12, 13). Используя протеомику, транскриптомику и экспериментальный анализ, Перна и его коллеги оценили несколько LILRB среди 24 лучших кандидатов-мишеней AML (14).LILRB действуют как молекулы иммунных контрольных точек и как факторы поддержания опухоли, но не могут влиять на нормальное развитие (8). Таким образом, у них есть потенциал в качестве привлекательных мишеней для лечения рака.
Моноцитарный ОМЛ представляет собой подтип ОМЛ, при котором большинство лейкозных клеток относятся к моноцитарной линии. Экстрамедуллярное заболевание, включая инфильтраты десен и поражение кожной и спинномозговой жидкости, часто встречается при моноцитарном ОМЛ (1). В соответствии с выводом Dobrowolska и коллег (15), мы сообщили, что LILRB4, член семейства LILRB, является маркером моноцитарного AML (16, 17).Мы также продемонстрировали, что LILRB4 более высоко экспрессируется на моноцитарных клетках AML, чем на их нормальных аналогах, и что экспрессия LILRB4 обратно коррелирует с общей выживаемостью пациентов с AML (16, 17). LILRB4 (также известный как CD85K, ILT3, LIR5 и HM18) имеет два внеклеточных Ig-подобных домена (D1 и D2) и три ITIM. Мы идентифицировали аполипопротеин E (ApoE) как внеклеточный связывающий белок LILRB4. Связывание ApoE связано с подавлением Т-клеток и инфильтрацией опухоли через LILRB4-опосредованную передачу сигналов ниже по течению в клетках AML (17).В совокупности эти данные показывают, что LILRB4 с рестриктивной и более низкой экспрессией на нормальных моноцитарных клетках является маркером моноцитарного AML с рестриктивной и более низкой экспрессией на нормальных моноцитарных клетках, который ингибирует активацию иммунной системы и поддерживает инвазивность опухоли. Следовательно, LILRB4 представляет собой привлекательную мишень для разработки лекарств для лечения пациентов с моноцитарным AML.
В этом исследовании мы сообщаем о гуманизированном mAb, направленном на LILRB4, h228-3, которое конкурентно блокирует взаимодействие LILRB4 / APOE.Это блокирующее антитело ингибирует инфильтрацию ткани моноцитарными клетками AML и отменяет подавление Т-клеток. Кроме того, h228-3 запускает ADCC- и ADCP-опосредованное уничтожение клеток AML. Обработка h228-3 значительно снизила опухолевую нагрузку AML на различных моделях мышей, включая PDX и сингенные иммунокомпетентные модели мышей. Эти результаты предполагают, что LILRB4-нейтрализующие антитела, такие как mAb h228-3, могут применяться в противораковых терапевтических стратегиях.
Материалы и методы
Клеточные линии и образцы AML человека
Клеточные линии HEK293F и CHO были получены от Life Technologies.Линии моноцитарных клеток AML человека (THP-1, MV4-11 и U937), линия клеток лейкемии мыши C1498 и линия клеток макрофагов мыши RAW264.7 были получены из АТСС и поддерживались в увлажненной атмосфере с 5% CO 2 ат. 37 ° C, в средах, предложенных ATCC с добавлением фетальной бычьей сыворотки (FBS; HyClone) и 100 Ед / мл пенициллина и 100 мкг / мл стрептомицина (Life Technologies). Клеточные линии не были аутентифицированы в прошлом году и культивировали менее 10 пассажей в указанной среде. Все клеточные линии обычно тестировали с использованием набора для контаминации Mycoplasma (R&D Systems).Первичные образцы ОМЛ человека были получены из Юго-западного медицинского центра Техасского университета (UTSW). Информированное согласие было получено в соответствии с протоколом, рассмотренным и утвержденным Советом по институциональной проверке UTSW. Образцы, экспрессированные LILRB4, анализировали с помощью проточной цитометрии.
Животные
Мыши C57BL / 6 и NOD-SCID с нулевым уровнем IL2Rγ (NSG) были приобретены и содержались в центре содержания животных UTSW. Для каждого эксперимента использовали мышей того же пола и того же возраста (4–8 недель), которые случайным образом распределяли по каждой группе.Все эксперименты на животных проводились с одобрения Институционального комитета по уходу и использованию животных UTSW.
Получение кроличьих mAb LILRB4
Двух новозеландских белых кроликов иммунизировали подкожно 0,5 мг рекомбинантно экспрессированного белка ECD человека LILRB4 (Sino Biological). После первоначальной иммунизации животным вводили ревакцинации четыре раза с интервалом в 3 недели. Титры сыворотки оценивали с помощью непрямого иммуноферментного анализа (ELISA), и В-клетки памяти выделяли после пятикратной иммунизации (RevMab Biosciences).Собирали большую панель одиночных В-клеток памяти и культивировали в течение 2 недель, а супернатанты анализировали с помощью ELISA (RevMab). Гены вариабельной области из этих положительных единичных В-клеток были выделены с помощью ПЦР с обратной транскрипцией (ОТ-ПЦР) с использованием праймеров (RIgH-F: ATGGAGACTGGGCTGCGCTGGCTYC; RIgH-R: CCATTGGTGAGGGTGCCCGAG; RIgK-F: ATGGCCGACCAGTAG; RIgK-F: ATGGCCGCCI были специфичны для вариабельных областей тяжелой и легкой цепи кролика. Два раунда ПЦР были выполнены путем включения перекрывающихся последовательностей на 3′- и 5′-концах, что позволило клонировать вариабельные области путем инфузии в векторы для экспрессии тяжелых и легких цепей кролика.Конструкции тяжелых и легких цепей котрансфицировали в клетки эмбриональной почки человека freestyle 293 (HEK293F) с использованием реагента для трансфекции PEI (Sigma; ссылка 18). После 7 дней экспрессии супернатанты собирали и антитела очищали аффинной хроматографией с использованием смолы с протеином А, как мы сообщали ранее (Repligen; ссылка 18). Панель из 26 очищенных кроличьих моноклональных антител (mAb) была создана и использована для этого исследования.
Анализ репортерных химерных рецепторов
Мы сконструировали стабильную систему репортерных клеток химерных рецепторов, как описано ранее (19).Одна из целей этой системы состояла в том, чтобы проверить способность лиганда связываться с ECD LILRB4. Другой целью системы было запустить активацию или ингибирование химерно слитого внутриклеточного домена парного иммуноглобулиноподобного рецептора (PILR) β. Этот рецептор передает сигнал через адаптер DAP-12 на активацию промотора NFAT. Если агонистическое антитело связывает ECD и активирует химерный сигнальный домен, наблюдается увеличение экспрессии GFP. Если антагонистическое антитело связывает ECD и подавляет химерный сигнальный домен, наблюдается снижение экспрессии GFP.Для скрининга антител, блокирующих LILRB4, использовали конкурентный анализ. Вкратце, рекомбинантный белок APOE (10 мкг / мл) был предварительно нанесен на 96-луночные планшеты при 37 ° C в течение 3 часов. После двух промывок PBS в каждую лунку засевали 2 × 10 4 репортерных клеток LILRB4. Тем временем указанные антитела LILRB4 добавляли в культуральную среду. После культивирования в течение 16 часов процент репортерных клеток GFP + измеряли с помощью проточной цитометрии.
Объединение эпитопов с помощью биослойной интерферометрии (BLI)
Система Octet RED96, биосенсоры протеина А и кинетический буфер были приобретены у ForteBio.Данные по объединению эпитопов были получены с помощью сэндвич-анализа по объединению эпитопов на основе BLI, выполненного на 8-канальном приборе Octet RED96. Сначала антитела (40 мкг / мл) загружали на биосенсоры протеина А в течение 4 минут. Оставшиеся сайты связывания Fc на биосенсорах блокировали нерелевантным кроличьим антителом (200 мкг / мл) в течение 4 минут с последующим замачиванием биосенсоров в кинетическом буфере на 10 секунд. Затем биосенсоры подвергали воздействию рекомбинантного LILRB4 (25 мкг / мл) в течение 4 минут для насыщения сайта связывания первого антитела.Наконец, биосенсоры подвергали воздействию вторичных антител (40 мкг / мл) в течение 4 минут для обнаружения связывания. Если не наблюдалось увеличения сигнала связывания со вторым антителом по сравнению с сигналом связывания первого антитела, пара антител была отнесена к одному и тому же эпитопному блоку (конкурент). Напротив, если наблюдалось усиление сигнала связывания со вторым антителом по сравнению с сигналом связывания первого антитела, пара антител была классифицирована в различных блоках эпитопов (неконкурентные).Бункеры — это группы антител, которые распознают один и тот же сайт / область связывания антигена. Биосенсоры с белком А использовали повторно 10 раз, и поверхности регенерировали в течение 30 секунд в 100 ммоль / л глицина (pH 2,6). Необработанные данные были обработаны с использованием программного обеспечения для анализа данных ForteBio 7.0.
Измерение аффинности с помощью BLI
Для измерения аффинности антитела антитело (30 мкг / мл) загружали в биосенсоры протеина G на 4 минуты. После короткого исходного уровня в кинетическом буфере загруженные биосенсоры подвергали воздействию серии рекомбинантных концентраций LILRB4 (0.1–200 нмоль / л) и вычитание фона использовалось для корректировки дрейфа датчика. Все эксперименты проводились при встряхивании 1000 об / мин. Сдвиги фоновых длин волн измеряли с помощью эталонных биосенсоров, которые были загружены только антителами. Программное обеспечение для анализа данных ForteBio использовалось для подгонки данных к модели связывания 1: 1 для извлечения скорости ассоциации и скорости диссоциации. K d было рассчитано с использованием отношения k off / k на .
Гуманизация кроличьего мАт
Гуманизация антитела LILRB4 была основана на стратегии трансплантации определяющих комплементарность областей (CDR), как описано ранее (20, 21). Вкратце, CDR в тяжелой и легкой цепях кроличьего антитела определяли с помощью комбинации трех методов: Kabat, IMGT и Paratome. Затем сравнивали родительские кроличьи mAb и наиболее близкую последовательность человеческой зародышевой линии. Остатки, которые, как известно, не имеют решающего значения для структуры и / или подвержены изменениям во время процесса созревания in vivo , были идентифицированы с помощью анализа на основе мутаций и гуманизированы (20).ДНК, кодирующие гуманизировать были синтезированы В. К. и В. (GenScript; ВЙ: EVQLLESGGGLVQPGGSLRLSCAASGIDFSNHYYMYWVRQAPGKGLEWIGSIFSGDSASTYYADSAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGMSTNDWASDLWGQGTLVTVSS; КИ: DIQMTQSPSSLSASVGDRVTITCQASESINSIYLAWYQQKPGKAPKLLIYRASTLASGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYDWGDVENTFGGGTKVEIK). Сигнальные пептиды человеческого IgG и последовательность Козака были сконструированы на 5′-концах последовательностей VK и VH. Затем гуманизированные фрагменты VK и VH клонировали по отдельности в векторы CK и CH человеческого IgG1.Экспрессия, очистка и количественная оценка гуманизированных mAb такие же, как и для кроличьих mAb.
Создание Fab D1 и h228-3 для работы структуры
ДНК, кодирующая домен D1 LILRB4, была клонирована в вектор pET21a (Novagen) с сайтами рестрикции Nde I и Xho I, а затем экспрессирована в E .coli , штамм BL21 (DE3; Novagen; ссылка 22). Бактерии культивировали в среде LB, содержащей соответствующие антибиотики (100 мкг / мл ампициллина), в шейкере-инкубаторе при 37 ° C.Экспрессию индуцировали добавлением 1 ммоль / л изопропил-β-D-1-тиогалактопиранозида (IPTG), когда культура достигла OD 600 0,8–1,0. Затем культивирование продолжали в течение 4-6 часов до сбора урожая. Центрифугированные клетки суспендировали в буфере PBS и разрушали с помощью гомогенизатора (JNBIO). Тельца включения рекомбинантных белков очищали и повторно укладывали, как описано, с некоторыми модификациями (22). Вкратце, аликвоты телец включения по каплям разводили в перемешивающем буфере для рефолдинга (100 ммоль / л Трис-HCl, 2 ммоль / л EDTA, 400 ммоль / л L-аргинин, 0.5 ммоль / л окисленного глутатиона и 5 ммоль / л восстановленного глутатиона, pH 8,0) в течение 8 часов при 4 ° C. Реорганизованный белок концентрировали и заменяли буфер с использованием концентратора Amicon 8400 на раствор, содержащий 20 ммоль / л трис-HCl и 50 ммоль / л NaCl, pH 8,0. Впоследствии белки дополнительно очищали гель-фильтрационной хроматографией. Подходящие фракционированные по пикам белки концентрировали для дальнейшего изучения или кристаллизации. Для получения Fab антитела концентрировали примерно до 20 мг / мл и расщепляли с использованием протеазы папаина (Pierce) при соотношении антитела к папаину 160: 1 (мас. / Мас.) При 37 ° C в течение 6 часов.Перевариваемую смесь загружали в колонку с протеином A (GE Healthcare), применяя проточный режим для разделения Fab-фрагмента с Fc-областью и непереваренным антителом. Fab-фрагменты собирали, концентрировали и очищали до гомогенности на колонке HiLoad 16/600 Superdex 200 пг (GE Healthcare).
Определение структуры комплекса h228-3-fab / D1
Белок D1 смешивали с очищенным Fab-фрагментом h228-3 в молярном соотношении 1,5: 1 и инкубировали при 4 ° C в течение ночи для образования комплекса.Затем смесь загружали в колонку Superdex 75 10/300 GL для очистки комплекса D1 / Fab от любого избытка D1. Пиковые фракции, соответствующие комплексу, собирали и концентрировали для кристаллизации. Кристаллы комплекса D1 / Fab были выращены диффузией пара в сидячих каплях. Всего 1 мкл раствора комплексного белка с концентрацией 5 мг / мл или 10 мг / мл смешивали с равным объемом резервуарного раствора. Кристаллы получали дифрагированное синхротронное излучение анизотропно с разрешением около 4,0 Å.Наконец, лучшее разрешение 3,0 Å было получено с использованием метода штрихового посева с собранными комплексными белками в том же резервуарном растворе при 4 ° C в течение 2 недель (PDB: 6K7O). Данные дифракции рентгеновских лучей были собраны при 100 K на канале синхротронного излучения в Шанхае BL17U и проиндексированы, интегрированы и масштабированы с помощью HKL2000. Сложная структура была решена методом молекулярной замены с использованием Phaser из пакета программ CCP4 со структурами LILRB4 (PDB: 3P2T) и Fab (PDB: 4OQT) в качестве моделей поиска.Первоначальная доработка фиксированного твердого тела была выполнена с использованием REFMAC5. Первоначальное ручное построение модели было выполнено с помощью COOT. Дальнейшее уточнение проводилось с помощью Phenix. Окончательная статистика для сбора данных и уточнения структуры представлена в дополнительной таблице S1.
Анализ проточной цитометрии
Для анализа приживления клеток AML человека у мышей NSG следовали ранее опубликованному протоколу (12). Клетки запускали либо на Calibur, либо на FACSAria для анализа. Исходные данные о потоке были проанализированы с помощью программного обеспечения FlowJo.Связывание mAb LILRB4 с клетками AML измеряли с использованием прибора Guava easyCyte HT в соответствии с инструкциями производителя (Millipore). Вкратце, 2 × 10 5 клеток распределяли в аликвотах по 100 мкл и блокировали 100 мкг / мл hIgG. Затем добавляли LILRB4 mAb (5 мкг / мл) в течение 1 часа на льду с последующим добавлением FITC-конъюгированного антикроличьего или античеловеческого вторичного IgG-F (ab) 2 (Jackson ImmuneResearch Laboratories). После промывания буфером PBS клетки анализировали на интенсивность флуоресценции.Нерелевантный кроличий или человеческий IgG использовали в качестве отрицательного контроля.
Получение мутантов LILRB4
Мутанты LILRB4 получали с использованием метода перекрытия двух циклов ПЦР с конструкцией ДНК LILRB4 дикого типа в качестве матрицы. Мутагенные праймеры были синтезированы Sigma (E54A-F: TGTCAGGGGACCCTGGCGGCTCGGGAGTACCGT; E54A-R: ACGGTACTCCCGAGCCGCCAGGGTCCCCTGACA; E54S-F: TGTCAGGGGACCCTGTCGGCTCGGGAGTACCGT; E54S-R: ACGGTACTCCCGAGCCGACAGGGTCCCCTGACA; E54R-F: TGTCAGGGGACCCTGAGGGCTCGGGAGTACCGT; E54R-R: ACGGTACTCCCGAGCCCTCAGGGTCCCCTGACA; R56E-F: GGGACCCTGGAGGCTGAGGAGTACCGTCTGGAT; R56E-R: ATCCAGACGGTACTCCTCAGCCTCCAGGGTCCC; R56A-F: GGGACCCTGGAGGCTGCGGAGTACCGTCTGGAT; R56A-R: ATCCAGACGGTACTCCGCAGCCTCCAGGGTCCC; R56Q-F: GGGACCCTGGAGGCTCAGGAGTACCGTCTGGAT; R56Q-Р: ATCCAGACGGTACTCCTGAGCCTCCAGGGTCCC; P103T-F: TGTTACTATCGCAGCACTGTAGGCTGGTCACAG; P103T-Р: CTGTGACCAGCCTACAGTGCTGCGATAGTAACA; P103R-F: TGTTACTATCGCAGCCGTGTAGGCTGGTCACAG; P103R- R: CTGTGACCAGCCTACACGGCTGCGATAGTAACA; P103S-F: TGTTACTATCGCAGCAGCGTAGGCTGGTCACAG; и P103S-R: CTGTGACCAGCCTACGCTGCTGCGATAGTAACA), и секвенирование ДНК было подтверждено с помощью GENEW.Затем фрагменты дикого типа и мутантные фрагменты клонировали в вектор с Fc-меткой hIgG1 с использованием метода клонирования вливания (Clontech). Эти мутанты LILRB4-Fc экспрессировали временной трансфекцией клеток HEK293F и очищали аффинной хроматографией на протеине А.
Выравнивание последовательностей и филогенетический анализ
Анализировали аминокислотные последовательности D1 11 членов семейства LILR. Номера доступа белков в GenBank следующие: LILRB1, Q8NHL6; LILRB2, Q8N423; LILRB3, O75022; LILRB4, Q8NHJ6; LILRB5, O75023; LILRA1, O75019; LILRA2, Q8N149; LILRA3, Q8N6C8; LILRA4, P59901; LILRA5, A6NI73; LILRA6, Q6PI73.Мы определили аминокислотные остатки от положения 27 до положения 118 как D1. Множественные выравнивания были выполнены с использованием ClustalX (версия 2.09) с 11 последовательностями D1. Филогенетическое дерево было создано с использованием MEGA (версия 5.0).
Анализ связывания ELISA
96-луночные планшеты Corning для EIA / RIA покрывали в течение ночи при 4 ° C рекомбинантными белками LILRB4 (1 мкг / мл) и блокировали в течение 2 часов при 37 ° C 5% обезжиренным молоком. После 3-кратной промывки PBST добавляли 100 мкл серийно разведенных антител LILRB4 и инкубировали в течение 45 минут при 37 ° C.Затем планшеты промывали PBST и инкубировали в течение 30 минут с IgG, конъюгированным с антителами против кролика или человека F (ab ‘) 2 HRP (Jackson ImmunoResearch Laboratories). Иммунореакции проявляли с субстратами TMB (Sigma) и останавливали добавлением 2 моль / л серной кислоты перед считыванием планшета при 450 нм.
Анализ апоптоза аннексином V / PI
Для анализа апоптоза, индуцированного антителом LILRB4, 5 × 10 4 клеток THP-1 высевали в 4 повторах в 12-луночные планшеты.Через 24 часа добавляли антитела до конечной концентрации 20 мкг / мл. Клетки обрабатывали нерелевантным человеческим IgG1 в качестве контроля изотипа. После 48 часов инкубации при 37 ° C в увлажненном воздухе с 5% CO 2 клетки собирали, дважды промывали PBS и ресуспендировали в 150 мкл связывающего буфера. К клеткам добавляли пять микролитров FITC-конъюгированного аннексина V и PI (йодид пропидия; BD Biosciences), встряхивали и инкубировали при комнатной температуре в темноте в течение 10 минут. Апоптоз измеряли с помощью проточной цитометрии.
Анализ совместного культивирования лейкозных клеток и Т-клеток
В анализе совместного культивирования человеческие Т-клетки (5 × 10 4 клеток на лунку), выделенные из периферической крови здорового донора (AllCells), помещали в нижние камеры 96- хорошо трансвелл-пластина. Клетки лейкемии культивировали в верхней камере вставок трансвелл (размер пор 3 мкм; Thermo Fisher) в 96-луночном планшете с U-образным дном. Облученные клетки указанной лейкемии (соотношение Е: Т = 2: 1) добавляли в верхние камеры и обрабатывали указанными антителами.После культивирования с гранулами, покрытыми анти-CD3 / CD28 (Thermo Fisher) и 50 ед / мл rhIL2 в нижних камерах в течение 5-7 дней, репрезентативные Т-клетки в нижних камерах фотографировали с использованием инвертированного микроскопа. Т-клетки окрашивали анти-CD3-APC, анти-CD4-PE или анти-CD8-PE и анализировали проточной цитометрией. Изотип мышиного IgG использовали в качестве отрицательного контроля. PI использовали в качестве индикатора мертвых клеток.
Анализ миграции через лунки
Для измерения миграционной способности клеток AML 2 × 10 5 клеток THP-1 высевали в верхнюю камеру (размер пор 8 мкм; Thermo Fisher) и обрабатывали антителами (10 мкг / мл).После культивирования в течение 20 часов клетки в нижней камере собирали и подсчитывали.
Хоуминг лейкозных клеток
Клетки (5 × 10 6 клеток на мышь) внутривенно вводили мышам NSG. Мышам вводили 10 мг / кг анти-LILRB4 или контрольного IgG1 сразу после инъекции лейкозных клеток. Мышей умерщвляли через 8 или 20 часов. Забирали периферическую кровь, костный мозг, печень и селезенку и исследовали суспензии одноклеточных клеток с помощью проточной цитометрии.CFSE, GFP или антитела против CD45 человека использовали для обнаружения клеток-мишеней лейкемии человека в указанных экспериментах. Количество лейкозных клеток в печени, селезенке и костном мозге реципиента указывается в виде отношения к количеству клеток в периферической крови. Для нормализации соотношение лейкозных клеток в разных органах, обработанных hIgG, нормализуется как 100%.
Ксенотрансплантат AML человека
Ксенотрансплантаты выполняли в основном, как описано (17, 23). Вкратце, для трансплантации использовали мышей NSG в возрасте от 6 до 8 недель.Клетки лейкемии человека ресуспендировали в 200 мкл PBS, содержащего 1% FBS. Мышам давали 1 × 10 6 культивируемых лейкозных клеток человека или 5-10 × 10 6 первичных клеток AML человека посредством инъекции в хвостовую вену. Через четыре часа после трансплантации вводили контрольный анти-LILRB4 или человеческий IgG1 путем инъекции в хвостовую вену. В указанные дни после трансплантации периферическую кровь, костный мозг, селезенку и печень оценивали на предмет приживления. Рост опухоли также отслеживали с течением времени по люминесценции.
Аллотрансплантат AML мыши
Аллотрансплантат AML мыши выполняли, как описано ранее (12, 17). Вкратце, для трансплантации использовали мышей C57BL / 6 дикого типа в возрасте от 6 до 8 недель. Клетки лейкемии мышей (1 × 10 6 ), экспрессирующие человеческий LILRB4, ресуспендировали в 200 мкл PBS для подкожной имплантации каждой мыши. Затем мышам вводили 10 мг / кг анти-LILRB4 или контрольного человеческого IgG внутривенно на 7 день после имплантации лейкозных клеток и обрабатывали каждые 3 дня до эвтаназии.Размеры опухоли определяли штангенциркулем (ширина × ширина × длина). Для истощения CD8 + Т-клеток 10 мг / кг антитела CD8 (YTS 169.4.2) вводили внутривенно на 3 день после имплантации лейкозных клеток и лечили дополнительно два раза каждые 3 дня. Чтобы определить, генерирует ли обработка антителом LILRB4 Т-клетки памяти, процентное содержание CD8 + CD62L + Т-клеток памяти в селезенке оценивали с помощью проточной цитометрии.
Анализ ADCC
Люминесцентная пленка человека была получена от здоровых доноров через Стэнфордский центр крови.Свежевыделенные PBMC человека (2,5 × 10 6 ) использовали в качестве эффекторных клеток, а 5 × 10 4 клеток THP-1 со стабильно трансфицированным GFP использовали в качестве клеток-мишеней в соотношении 50: 1. Человеческие PBMC, клетки-мишени THP-1-GFP и возрастающие концентрации антител были объединены в 200 мкл всего в RPMI + 10% инактивированной нагреванием FBS и 50 нг / мл IL2 (R&D Systems) в U-образном 96-луночном планшете. и инкубировали 20 часов при 37 ° C. Клетки промывали и ресуспендировали в 200 мкл окрашивающего буфера (BD Biosciences), содержащего 7-AAD.Клетки (150 000) были приобретены FACSCelesta, и было измерено процентное содержание GFP-положительных клеток. Цитотоксичность клеток рассчитывали как: процент цитотоксичности = 100 — ([T / NT] × 100), где T и NT — это процентное соотношение клеток GFP + , обработанных антителами или без них, соответственно.
Анализ CDC
Клетки THP-1 высевали в трех экземплярах в 96-луночный планшет с плотностью 2 × 10 4 клеток на лунку. Добавляли контроль изотипа анти-LILRB4 или человеческого IgG1 до конечной концентрации 20 мкг / мл.Через 30 минут добавляли нормальную человеческую сыворотку (Innovative) до конечной концентрации 20%. После инкубации в течение 4 часов при 37 ° C клеточные супернатанты переносили в 96-луночный планшет для определения количества высвобожденного ЛДГ с использованием набора для анализа цитотоксичности ЛДГ (Pierce).
ADCP анализ
In vitro антителозависимый клеточный фагоцитоз (ADCP) выполняли с клеточной линией макрофагов мыши RAW 264.7 (мыши Mac) или макрофагами, полученными из PBMC человека (Mac Mac), и анализировали с помощью проточной цитометрии (24).Клетки RAW 264.7, стимулированные липополисахаридом (LPS), или человеческие макрофаги окрашивали efluor 670 (красный) и высевали с плотностью 4 × 10 4 клеток на лунку и оставляли на ночь. Клетки-мишени THP-1 окрашивали CellVue Jade (зеленый), предварительно инкубировали с 20 мкг / мл контрольного изотипа анти-LILRB4 или человеческого IgG1 в течение 30 минут и добавляли в лунки в соотношении Е / Т 1: 1 в трех повторностях. После инкубации в течение 2-4 часов при 37 ° C суспензионные клетки THP-1 смывали PBS и прилипшие макрофаги мыши или макрофаги человека собирали для анализа проточной цитометрией.Процент ADCP рассчитывали по двойным положительным макрофагам / общему количеству макрофагов.
Статистический анализ
Статистический анализ выполняли с помощью Prism 7.0 (программное обеспечение GraphPad). Статистические различия были определены как значимые при P <0,05 с использованием двустороннего критерия Стьюдента t и двустороннего лог-рангового критерия Манна-Уитни. In vitro данные были представлены как среднее ± стандартная ошибка среднего. На всех рисунках: *, P <0,05; **, P <0.01.
Результаты
Получение и характеристика mAb LILRB4
Чтобы охарактеризовать потенциал блокады LILRB4 / APOE в качестве терапевтической стратегии для AML, мы создали панель mAb LILRB4. Протокол для создания этих mAb проиллюстрирован на фиг. 1A (20, 25, 26). Вкратце, мы серийно иммунизировали кроликов белком внеклеточного домена (ECD) LILRB4. Кролика, который показал наивысший сывороточный титр связывания LILRB4 в ELISA, отбирали для выделения и культивирования одиночных В-клеток памяти.Из 400 подвергнутых скринингу LILRB4-специфичных единичных клонов B-клеток, всего было получено 229 связывающих LILRB4 (дополнительные рисунки S1A и S1B). Затем мы клонировали вариабельные области последовательностей тяжелых и легких цепей из 26 лучших совпадений с наивысшей активностью связывания. После рекомбинантной экспрессии в клетках HEK293F в виде кроличьего IgG1 мы определили связывающую способность с помощью ELISA и обнаружили EC 50 для 21 антитела в низком наномолярном диапазоне (0,05–0,3 нмоль / л; рис. 1B). Чтобы сгруппировать mAb по их связывающим эпитопам, мы выполнили сэндвич-анализ связывания эпитопов с Octet RED96 (27).Всего для 21 высокоаффинного связывающего агента было идентифицировано семь эпитопных бункеров (от 1 до 7) (фиг. 1C). Чтобы дополнительно охарактеризовать связывающие эпитопы в каждом бункере, мы взяли репрезентативные антитела из каждого бункера и использовали ELISA для измерения их связывания с различными доменами LILRB4 ECD: D1 (остатки 27–118), D2 (остатки 119–218) и SR ( область стебля, остатки 219–259). Мы использовали полноразмерный LILRB4 ECD (остатки 22–259) в качестве контроля (дополнительный рис. S2). MAb в трех блоках эпитопа связываются с D1, mAb в одном блоке эпитопа — с D2, а mAb в трех блоках эпитопа — с SR (фиг.1D). Как и ожидалось, в анализе проточной цитометрии mAb из всех эпитопных связок связываются с клетками THP-1, моноцитарной линией клеток AML, экспрессирующей высокую плотность LILRB4. Этот результат предполагает, что эти mAb могут связываться с моноцитарными клетками AML в физиологических условиях (рис. 1E).
Рисунок 1.Получение и характеристика LILRB4 блокирующих mAb. A, Стратегия выделения, культивирования и клонирования единичных антиген-специфических В-клеток памяти, используемых для получения кроличьих mAb LILRB4. После выделения и культивирования LILRB4-специфических В-клеток памяти от иммунизированных кроликов желаемые В-клетки подвергали скринингу на связывание с LILRB4 в ELISA.Затем гены vH и vL клонировали в остова кроличьего IgG, и рекомбинантные mAb получали с использованием системы временной экспрессии клеток HEK293F. B, EC 50 из 26 кроличьих mAb LILRB4. Нерелевантное кроличье антитело (rIgG) использовали в качестве отрицательного контроля. EC 50 ≥ 1,0 нмоль / л отображается как 1,0 нмоль / л. Было проведено два независимых эксперимента. C, Узловой график бункеров эпитопов 21 LILRB4 кроличьих mAb, определенных Octet RED96 с использованием классического сэндвич-анализа биннинга эпитопов. D, Связывание семи репрезентативных кроличьих mAb (из разных бункеров) с ECD (остатки 22–259), D1 (первый Ig-подобный домен, остатки 27–118), D2 (второй Ig-подобный домен, остатки 119–218). ) и SR (участок стебля, остатки 219–259) LILRB4, определенный с помощью ELISA. E, Связывание семи репрезентативных кроличьих mAb (из разных бункеров) с LILRB4-экспрессирующими клетками THP-1 определено с помощью проточной цитометрии. Было проведено два независимых эксперимента. F, Скрининг mAb, блокирующих LILRB4, в анализе репортерного химерного рецептора.APOE2 использовали в качестве функционального лиганда для активации репортерных клеток LILRB4. Два блокирующих антитела LILRB4 показаны красным. G, и H, Кинетику связывания 128-3 и 216-1 с LILRB4 оценивали с использованием Octet RED96. I, Связывающая способность 128-3 и 216-1 с LILRB4 определена с помощью ELISA. J, LILRB4 эффективность блокирования 128-3 и 216-1 определяли репортерным анализом. Было проведено два независимых эксперимента. K, Специфичность 128-3 и 216-1 оценивали с помощью ELISA. L, Комбинированную стратегию трансплантации CDR KABAT / IMGT использовали для гуманизации кроличьих mAb 128-3. M, Связывание гуманизированного 128-3 (h228-3) с LILRB4 определяли с помощью ELISA. N, Сродство гуманизированного 128-3 (h228-3) к LILRB4 определяли с помощью Octet RED96. Эффективность блокирования h228-3 O, LILRB4 определяли репортерным анализом. Было проведено два независимых эксперимента.
Идентификация и гуманизация мАт, блокирующего LILRB4
Чтобы проверить, блокируют ли эти мАт LILRB4 активацию LILRB4 с помощью APOE, функционального внеклеточного связывающего белка LILRB4, мы проверили 21 мАт с использованием репортерного анализа химерного рецептора LILRB4 на основе GFP (дополнительный рис. .S3; исх. 17). Было обнаружено, что два антитела из одного и того же эпитопного бункера (блок 4) нейтрализуют APOE-опосредованную активацию LILRB4 (рис. 1F). Используя Octet RED96, мы дополнительно оценили кинетику связывания LILRB4 этих двух mAb, 128-3 и 216-1. Значения Kd для связывания с LILRB4 составляют 2,9 нмоль / л для 128-3 и 14,5 нмоль / л для 216-1 (рис. 1G и H). Мы выбрали 128-3 в качестве ведущего mAb на основании его более высокой связывающей активности LILRB4, измеренной с помощью ELISA (рис. 1I), более высокого сродства к LILRB4, измеренного системой Octet RED96 (рис.1G и H), а также более сильную эффективность блокирования LILRB4 / APOE, как оценивали в анализе репортерного химерного рецептора (фиг. 1J). Неспецифическое связывание терапевтического антитела может вызвать серьезные побочные эффекты из-за активности, не связанной с мишенью. Семейство рецепторов LILR содержит 11 членов (B1-B5 и A1-A6), которые тесно связаны филогенетически, особенно в домене D1 (дополнительный рисунок S4). Эти рецепторы экспрессируются на широком спектре нормальных клеток. Чтобы определить специфичность 128-3, мы затем экспрессировали полноразмерные белки ECD для всех 11 рецепторов семейства LILR и проверили их связывание с 128-3 с помощью ELISA.Как показано на фиг. 1K, mAb 128-3 специфично для LILRB4 и не реагирует перекрестно с другими членами семейства LILR.
Для потенциальной терапевтической разработки мы использовали стратегию прививки CDR для гуманизации 128-3 (h228-3), превращая его в подкласс человеческого IgG1 с легкой цепью каппа (рис. 1L). Затем мы измерили аффинность связывания h228-3 с LILRB4 с помощью Octet RED96 и ELISA. Мы также измерили эффективность блокирования LILRB4 с помощью анализа репортерных химерных рецепторов. ЕС 50 для связывания h228-3 с LILRB4, измеренный в ELISA, равен 0.23 нмоль / л. Это менее чем в три раза превышает ЕС 50 0,08 нмоль / л для родительского 128-3 (фиг. 1M). Точно так же Kd связывания h228-3 с LILRB4, измеренная с помощью Octet RED96, 3,5 нмоль / л, сравнима с Kd родительского 128-3, 2,9 нмоль / л (рис. 1N). IC 50 для активности блокирования LILRB4 для гуманизированного h228-3 составляет 0,24 нмоль / л, а IC 50 для родительского 128-3 составляет 0,14 нмоль / л. Эти результаты предполагают, что h228-3 сохранял аффинность связывания и блокирующую активность родительского кроличьего mAb 128-3 (фиг.1М – О).
Структурное выяснение взаимодействия h228-3 / LILRB4
Исследования связывания между h228-3 и различными доменами ECD LILRB4 показали, что антитело связывается с доменом D1 LILRB4 (рис. 2A). Затем мы попытались использовать рентгеновскую кристаллографию, чтобы выяснить молекулярную основу этого взаимодействия и охарактеризовать эпитоп на LILRB4, способствующий связыванию h228-3. Для достижения этой цели мы экспрессировали домен D1 LILRB4 в виде телец включения в E. coli .Из этих телец включения мы получили растворимые белки путем рефолдинга in vitro . Затем Fab-комплекс D1 / h228-3 готовили для скрининга кристаллов. Структура комплекса была определена путем молекулярного замещения с разрешением 3,0 Å (дополнительная таблица S1). Общая структура показывает, что h228-3 связывается с кончиком домена D1 (Fig. 2B). MAb h228-3 использует как тяжелую (VH), так и легкую (VL) цепи для взаимодействия с LILRB4, включая пять петель CDR во всех областях VH и VL h228-3, кроме LCDR2 (рис.2C и D; Дополнительная таблица S2). В частности, h228-3 связывается с петлями BC, C’E и FG D1. Остаток E54 в петле BC D1 образует четыре водородные связи с остатками как в областях HCDR1 (Y34), так и HCDR2 (S59 и Y61) (рис. 2C). Остаток R56 в петле BC взаимодействует с остатком Y34 в HCDR1, образуя одну водородную связь (рис. 2C). Кроме того, R56 вносит две водородные связи с остатками Y33 в LCDR1 и S92 в LCDR3 (рис. 2D). Кроме того, E76 и K78 в петле C’E D1 контактируют с S103 и D106 в HCDR3, образуя две водородные связи (рис.2С). Остатки R101 и P103 в петле FG D1 образуют пять водородных связей с остатками Y93, D94 и W95 в LCDR3 (рис. 2D). Поверхность связывания в D1 LILRB4 показана на фиг. 2E. Таким образом, три петли D1 нацелены на h228-3 через серию контактов, включая множественные взаимодействия водородных связей.
Рисунок 2.Расшифровка взаимодействия h228-3 с D1. A, Связывание h228-3 с D1, D2 и ECD LILRB4 определяли с помощью ELISA. B, Общая кристаллическая структура комплекса D1 / h228–3-Fab.D1 отображается серым цветом; тяжелая (H) цепь антитела h228-3 показана голубым, а его легкая (L) цепь — розовым. Цикл FG D1 показан желтым, цикл BC D1 показан зеленым, а цикл C’E D1 показан синим. C, Подробное описание взаимодействия D1 / h228–3-VH. D, Подробное описание взаимодействия D1 / h228–3-VL. Остатки, участвующие во взаимодействии водородных связей, показаны в виде палочек и помечены. Водородные связи показаны красными пунктирными линиями. E, Остатки эпитопа в D1 помечены черными символами.Остатки, с которыми контактирует h228-3-Fab VH, окрашены в голубой цвет. Остатки, с которыми контактирует h228-3-Fab VL, окрашены в розовый цвет. Остатки, с которыми контактируют обе цепи, окрашены в красный цвет. Связывание мутантов тяжелой ( F ) и легкой ( G ) цепи h228-3 с LILRB4 проводили с помощью ELISA. H, Получение и очистка мутантов D1, которые были слиты с меткой Fc человеческого IgG1 и экспрессированы в клетках HEK293F. I, Связывание h228-3 с мутантами D1 определяли с помощью ELISA. J, Связывание h228-3 с мутантами LILRB4 ECD.Три идентифицированных критических аминокислотных остатка, которые были мутированы на три различных типа аминокислотных остатков. K, Выравнивание последовательностей связывающего мотива h228-3 (петля BC, петля C’E и петля FG) во всех 11 членах семейства LILR. Уникальные аминокислотные остатки LILRB4 R56, R101 и V104 отмечены зеленым. Показаны общие аминокислотные сходства D1 в процентах по сравнению с LILRB4 (100%).
Чтобы подтвердить роль этих ключевых остатков в связывании LILRB4, мы создали одиночные аланиновые мутанты h228-3 (Y34A в HCDR1, S103A в HCDR3, Y33A в LCDR1 и Y93A в LCDR3).Мутантные антитела тестировали с помощью ELISA для оценки влияния на связывание LILRB4. Одиночные мутантные варианты Y34A, Y33A и Y93A полностью отменяли связывание h228-3 с LILRB4. Только S103A сохранил частичную связывающую способность LILRB4 (фиг. 2F и G). Мы также создали мутанты с одной аминокислотой в домене LILRB4 D1: E54A, R56Q и P103S. Среди членов семейства LILRB LILRB3 является ближайшим родственником LILRB4 на основании сходства аминокислот D1. Следовательно, мутации были разработаны на основе выравнивания последовательностей доменов D1 LILRB4 и LILRB3.Если остатки различаются между LILRB4 и LILRB3, аминокислотный остаток LILRB4 был заменен соответствующим остатком в LILRB3. Если остаток является консервативным в LILRB4 и LILRB3, он был изменен на аланин. В соответствии с предсказаниями анализа кристаллической структуры комплекса D1 / h228-3 Fab, все три мутации в домене LILRB4 D1 отменяют связывание с h228-3 (Fig. 2H-J).
Выравнивание связывающего мотива h228-3 (петля BC, петля C’E и петля FG) во всех 11 членах семейства LILR показало, что этот мотив является уникальным для LILRB4: R56, R101 и V104 являются LILRB4-специфичными аминокислотами остатки (рис.2К). Как и ожидалось, учитывая отсутствие перекрестной реактивности 128-3 с другими членами семейства LILR (фиг. 1K), результаты анализа кристаллической структуры и картирования эпитопа дополнительно подтвердили, что h228-3 является LILRB4-специфическим mAb. В соответствии с выводами о том, что петля FG является критической для активации LILRB4, индуцированной APOE (17), наши результаты показывают, что h228-3 блокирует ApoE-индуцированную активацию передачи сигналов LILRB4 конкурентным образом.
Отмена супрессии Т-клеток с помощью h228-3
Ранее мы продемонстрировали, что LILRB4, экспрессируемый на моноцитарных клетках AML, подавляет опосредованный Т-клетками противоопухолевый иммунитет через APOE / LILRB4 / SHP-2 / NF-κB / uPAR / Arginase- 1 ось (17).Аргиназа-1 является ключевым последующим эффектором передачи сигналов LILRB4 / NF-kB / uPAR и может секретироваться клетками AML для подавления активности Т-клеток. В этом исследовании мы стремились определить, отменяет ли блокирование передачи сигналов LILRB4 с помощью mAb h228-3 супрессию Т-клеток. Мы совместно культивировали Т-клетки человека с клетками THP-1 и обрабатывали их h228-3. hIgG служил контролем. Обработка h228-3 значительно увеличивала пролиферацию Т-клеток в этих исследованиях совместно культивированных Т-клеток и клеток THP-1 (фиг. 3A). Количество CD3 + , CD4 + и CD8 + Т-клеток было выше в сокультурах, обработанных mAb h228-3 (рис.3B – D). Этот результат предполагает, что способность клеток THP-1 подавлять Т-клетки была отменена блокадой LILRB4. Более того, в анализе совместного культивирования клеток PBMC и AML THP-1 обработка h228-3 увеличивала секрецию множества цитокинов, связанных с пролиферацией и активацией Т-клеток, включая CXCL9, CXCL11, IFNγ, IL2 и IL7 (рис. 3E). ).
Фигура 3.h228-3 отменяет подавление Т-клеток in vitro и in vivo . A, Репрезентативное изображение Т-клеток в анализе сокультивирования.Т-клетки, выделенные от здоровых доноров, инкубировали в нижней камере 96-луночного транслуночного планшета с облученными клетками THP-1 (E: T 2: 1) в верхней камере, разделенной мембраной с порами 3 мкм. После совместного культивирования с гранулами, покрытыми анти-CD3 / CD28, и rhIL2 в течение 7 дней репрезентативные клетки фотографировали с использованием инвертированного микроскопа (масштабная линейка, 100 мкм). Было проведено два независимых эксперимента. Т-клетки окрашивали анти-CD3 ( B ), анти-CD4 ( C ) и анти-CD8 ( D ) и анализировали проточной цитометрией. E, Количественный анализ цитокинов в супернатантах PBMC человека и анализ сокультивирования клеток THP-1. Человеческие PBMC (1,5 × 10 6 клеток / мл) и клетки THP-1 (3 × 10 5 клеток / мл) вместе с h228-3 или hIgG (20 мкг / мл) совместно культивировали в течение 48 часов. Затем супернатанты собирали и детектировали с использованием набора RayBio G-Series человеческих цитокиновых антител 1000 Kit. F, Дизайн исследования. Мышам C57BL / 6 подкожно имплантировали человеческие LILRB4, принудительно экспрессирующие мышиные клетки AML C1498 (C1498-hlilrb4) с последующей обработкой h228-3, h228-3 вместе с анти-CD8, контрольным человеческим IgG (hIgG) или hIgG вместе с анти- CD8.Конечные точки оценивали на 27 день после трансплантации клеток AML. G, Рост опухоли у мышей, несущих подкожно C1498-hlilrb4 ( n = 5), получавших h228-3 или hIgG. H, Количественное определение CD8 + CD62L + Т-клеток памяти у мышей, несущих подкожно C1498-hlilrb4, получавших h228-3 или hIgG. I, Рост опухоли подкожно несущих C1498-hlilrb4 мышей ( n = 5), получавших h228-3 или hIgG в состоянии истощения Т-клеток.
Чтобы дополнительно оценить, отменяет ли h228-3 супрессию Т-клеток in vivo , мы затем создали человеческую линию клеток AML, экспрессирующую LILRB4 человека, C1498-hlilrb4. Мы подкожно имплантировали клетки C1498-hlilrb4 мышам C57BL / 6 для создания сингенной иммунокомпетентной модели мыши. В этой модели обработка 10 мг / кг h228-3 на 6 день после имплантации клеток C1498-hlilrb4 значительно снизила опухолевую нагрузку и увеличила процент CD8 + CD62L + Т-клеток памяти в селезенке, что позволяет предположить h228-3 отменил подавление Т-клеток in vivo (рис.3F – H). Истощение CD8 + Т-клеток в этой модели мышей обработкой антителами против CD8 почти полностью устраняет противоопухолевую эффективность h228-3 (фиг. 3I). Вместе эти результаты предполагают, что h228-3 восстанавливает активность Т-клеток против опухолей in vitro, и in vivo, .
Блокирование тканевой инфильтрации и индукция мобилизации клеток AML с помощью mAb h228-3
Одной из характерных черт моноцитарного AML является усиленная экстрамедуллярная инфильтрация опухолевых клеток (28).Как мы сообщали ранее, LILRB4 направляет лейкозные клетки к миграции во внутренние органы через ось APOE / LILRB4 / SHP-2 / NF-κB / uPAR / Arginase-1 (17). Истощение LILRB4 в клетках AML значительно снижает их способность к хомингу in vivo (фиг. 4A). Чтобы определить, ингибирует ли h228-3 миграцию клеток AML, мы провели анализ миграции через лунки с использованием клеток AML THP-1. Обработка h228-3 значительно уменьшала миграцию клеток AML in vitro (фиг. 4B). Для дальнейшего изучения того, ингибирует ли h228-3 инфильтрацию ткани клеток AML in vivo , мы оценили его эффективность в отношении блокирования самонаведения клеток AML на модели NSG мышей.Клетки AML MV4-11 внутривенно вводили мышам NSG с последующей немедленной обработкой h228-3 или hIgG в качестве контроля. Обработка h228-3 значительно уменьшала кратковременное (8 часов и 20 часов) возвращение клеток AML MV4-11 в костный мозг (BM), печень (LV) и селезенку (SP; фиг. 4C и D). Более того, h228-3-опосредованная блокада LILRB4 показала значительное ингибирование долговременной (21 день) тканевой инфильтрации клеток AML MV4-11 в костный мозг, печень и селезенку (рис. 4E). В соответствии с h228-3, блокирующим самонаведение и тканевую инфильтрацию клеток AML MV4-11, биолюминесцентная визуализация всего животного показала, что обработка h228-3 значительно блокировала образование клеток AML THP-1, замедляла потерю массы тела и увеличивала выживаемость ксенотрансплантатов NSG. мышей (рис.4F – I). Чтобы оценить, является ли блокирование AML h228-3 у мышей NSG дозозависимым, клетки THP-1, экспрессирующие люциферазу (THP-1-luc), вводили внутривенно мышам NSG с последующим в тот же день обработкой однократной дозой h228-3 в разных дозах (0,01, 0,1 и 1 мг / кг). Визуализация биолюминесценции показала, что h228-3 блокирует приживление клеток AML THP-1 у мышей NSG дозозависимым образом. Лечение 1 мг / кг h228-3 показало гораздо более высокую терапевтическую активность, чем лечение 0.01 или 0,1 мг / кг (рис. 4J). Мы также наблюдали временную зависимость блокирования h228-3 тканевой инфильтрации клетками AML THP-1 тканевой инфильтрацией. Обработка h228-3 в день 0 после инъекции клеток AML показала лучшую эффективность в снижении инфильтрации ткани клетками AML у мышей NSG. Лечение на 12-й день после инъекции клеток AML оказало минимальное влияние на инфильтрацию клеточной ткани AML (дополнительный рисунок S5).
Рисунок 4.h228-3 блокирует миграцию клеток AML и тканевую инфильтрацию. A, Сравнение краткосрочной (20 часов) инфильтрации ткани клетками WT или lilrb4 -KO AML MV4-11 у мышей NSG ( n = 4).Количество лейкозных клеток (GFP + ) в BM, печени (LV) и селезенке (SP) определяли с помощью проточной цитометрии и нормализовали до количества в PB. Коэффициент самонаведения клеток MV4-11 у мышей, получавших hIgG, был нормализован до 100%. B, Сравнение способности клеток THP-1 к миграции через лунки, обработанных h228-3 или hIgG. Было проведено два независимых эксперимента. Кратковременные возможности самонаведения меченных CFSE клеток MV4-11, которые вводили мышам NSG с последующей немедленной обработкой h228-3 или hIgG через 8 ( C ) или 20 ( D ) часов после инъекции. E, Длительная (21 день) инфильтрация ткани клетками THP-1 у мышей NSG после обработки h228-3 или hIgG. F, Дизайн исследования. Мышам NSG ( n = 5) внутривенно вводили клетки THP-1, экспрессирующие люциферазу (THP-1-luc), с последующей немедленной обработкой h228-3 или hIgG. Биолюминесцентная визуализация ( G ), кривая выживаемости ( H ) и изменения массы тела ( I ) мышей NSG, получавших h228-3 или hIgG. Мышам J, NSG инъецировали 1 × 10 6 клеток THP-1-luc с последующей обработкой 0.01 мг / кг, 0,1 мг / кг или 1 мг / кг h228-3 и контролировали с помощью биолюминесцентного изображения. К, Дизайн исследования. Первичные клетки AML от пациентов с моноцитарным AML вводили облученным мышам NSG. После 9 недель приживления мышей умерщвляли и ткани (суспензию одноклеточных) с привитыми клетками AML затем инъецировали другим облученным мышам NSG с последующей немедленной обработкой h228-3 или hIgG. Конечные точки оценивали на 21 день после инъекции клеток AML. Клетки AML человека в BM, печени, селезенке и PB анализировали с помощью проточной цитометрии. L, Процент человеческих клеток AML (CD45 + LILRB4 + ), привитых в указанные органы, анализировали с помощью проточной цитометрии на 21 день после трансплантации. М, Дизайн исследования. Мышам NSG ( n = 5 или 6) внутривенно вводили клетки THP-1 с последующей обработкой цитарабином (10 мг / кг) и h228-3 или hIgG (10 мг / кг) в указанные моменты времени. Конечные точки оценивали на 21 день после инъекции клеток THP-1. Анти-человеческий CD45 использовали для обнаружения клеток лейкемии человека (клетки THP-1) в печени с помощью проточной цитометрии. N, Показаны проценты человеческих клеток, привитых в печень на 21 день после трансплантации.
Затем мы попытались оценить, можно ли воспроизвести эффекты блокирования тканевой инфильтрации h228-3 в модели мыши NSG ксенотрансплантата клеток AML THP-1 с первичными человеческими моноцитарными клетками AML. С этой целью мы впервые получили первичные клетки ОМЛ от пациентов с моноцитарным ОМЛ (Юго-западный медицинский центр Юта). Затем мы внутривенно трансплантировали эти клетки облученным мышам NSG на 9 недель.Ткани мыши вместе с привитыми человеческими клетками AML затем инъецировали другим облученным мышам NSG с последующей обработкой h228-3 или hIgG. Процентное содержание человеческих CD45 + LILRB4 + AML-клеток в костном мозге, печени, селезенке и периферической крови (PB) инъецированных мышей NSG измеряли с помощью проточной цитометрии (фиг. 4К и L). Обработка h228-3 значительно ингибировала развитие AML в первичных моноцитарных ксенотрансплантатах AML человека (фиг. 4L). В совокупности эти результаты показали, что h228-3 ингибирует миграцию моноцитарных клеток AML и тканевую инфильтрацию in vitro и in vivo .
Химиотерапия имеет ограниченную способность воздействовать на раковые клетки в нишах. Основываясь на способности LILRB4 поддерживать хоминг, мы выдвигаем гипотезу, что блокирование передачи сигналов LILRB4 усиливает мобилизацию этих размещенных в нишах клеток AML в циркуляцию. Если это так, то антитело, блокирующее LILRB4, повысит эффективность химиотоксических препаратов против AML, которые убивают опухолевые клетки за пределами их ниш. Чтобы проверить эту гипотезу, мы выполнили комбинацию h228-3 и цитарабина на мышиной модели NSG с ксенотрансплантатом клеток THP-1.После трансплантации клеток THP-1 мышам NSG, мышей обрабатывали цитарабином вместе с h228-3 или hIgG на 6 или 14 день (фиг. 4M). Блокирование LILRB4 с помощью h228-3 значительно усиливает анти-AML эффекты цитарабина (фиг. 4N). Эти результаты предполагают, что h228-3 может быть дополнительно разработан как часть комбинированной терапии с химиотерапевтическими препаратами для лечения ОМЛ.
ADCC и ADCP, опосредованные mAb h228-3, способствуют уничтожению опухолевых клеток
Fc-опосредованные иммунные функции, такие как антителозависимая клеточная цитотоксичность (ADCC), ADCP и комплемент-зависимая цитотоксичность (CDC), были утверждены как способы действие терапевтических мАт, используемых в терапии рака (29, 30).h228-3 не влияет на апоптоз или пролиферацию клеток AML THP-1 (фиг. 5A и B). Для дальнейшего исследования возможных Fc-опосредованных механизмов анти-AML эффектов in vitro, и in vivo , h228-3 анализировали на активность CDC, ADCC и ADCP с использованием клеток комплемента или эффекторных клеток. В эксперименте с использованием клеток THP-1 в качестве клеток-мишеней и нормальной человеческой сыворотки в качестве комплемента не было обнаружено активности CDC h228-3 даже при высокой концентрации антител 20 мкг / мл (фиг. 5C). Мы обнаружили активность ADCC и ADCP для h228-3.Клетки AML THP-1 были специфически убиты PBMC человека в присутствии h228-3 дозозависимым образом со значением EC 50 , равным 13,5 нмоль / л (фиг. 5D). Более того, h228-3 может направлять как макрофаги мыши (мышиные Mac, клетки RAW 264.7, стимулированные LPS), так и макрофаги, происходящие из PBMC человека (Macs человека, дифференцированные из моноцитов CD14 + , выбранных из PBMC человека), на фагоцитоз AML THP-1 клеток. в тесте ADCP проточной цитометрии (рис. 5E – G). Чтобы проверить, что Fc-опосредованная иммунная функция является одним из механизмов анти-AML активности h228-3, мы создали Fc-мутированный h228-3 (h228-3-N297A), который является дефектным для ADCC и ADCP, устраняя связывание h228-3 с FcγR на иммунных клетках (31).Как и ожидалось, h228-3 и h228-3-N297A показали такую же аффинность связывания с LILRB4, как определено с помощью ELISA (фиг. 5H). Затем мы попытались определить вклад h228-3 Fc-опосредованной активности против AML в гибель лейкозных клеток in vivo . Для достижения этой цели мы оценили активность h228-3 и h228-3-N297A у мышей NSG с ксенотрансплантатами клеток AML THP-1 (рис. 5I). Обработка 10 мг / кг h228-3-N297A коррелировала со значительно более высоким приживлением клеток AML THP-1 у мышей NSG, чем обработка h228-3 (рис.5J). Взятые вместе, эти результаты предполагают, что опосредованные h228-3 Fc иммунные функции вносят вклад в активность против AML in vitro и in vivo .
Рисунок 5.h228-3 запускает ADCC и ADCP. A, Сравнение апоптоза клеток THP-1, индуцированного h228-3 или hIgG. Было проведено два независимых эксперимента. B, Сравнение влияния на пролиферацию клеток THP-1, обработанных h228-3 или hIgG. Было проведено два независимых эксперимента. C, CDC h228-3 оценивали в анализе лактатдегидрогеназы (LDH) с использованием нормальной человеческой сыворотки в качестве комплемента и клетки THP-1 в качестве клетки-мишени.Было проведено два независимых эксперимента. D, ADCC, запускаемый h228-3 или hIgG, оценивали с помощью проточной цитометрии. Клетки THP-1-GFP использовали в качестве клеток-мишеней, а свежие изолированные PBMC от здоровых доноров использовали в качестве эффекторных клеток в соотношении E: T 50: 1. Было проведено два независимых эксперимента. E – G, ADCP, запускаемый h228-3 или hIgG, был обнаружен в анализе проточной цитометрии с использованием клеток THP-1 в качестве клеток-мишеней и линии клеток макрофагов мыши RAW 264.7 (мыши Mac) или макрофагов, полученных из PBMC человека (Mac Mac), как эффекторные клетки.После инкубации h228-3 или hIgG вместе с клетками-мишенями и эффекторными клетками в течение 2 часов при 37 ° C прилипшие макрофаги собирали и определяли проточной цитометрией. Процент фагоцитоза рассчитывали по двойным положительным макрофагам / общему количеству макрофагов. Показаны типичные профили FACS. Было проведено три независимых эксперимента. H, Сравнение связывания h228-3 дикого типа (h228-3) и N297A-мутантного h228-3 (h228-3-N297A) с LILRB4 в ELISA. I, Дизайн исследования. Мышам NSG ( n = 5) внутривенно вводили клетки AML THP-1 с последующим лечением h228-3 или h228-3-N297A на 3 день после инъекции клеток THP-1.Конечные точки оценивали на 21 день после инъекции клеток THP-1. Клетки человека (клетки THP-1) в BM, печени, селезенке и PB анализировали с помощью проточной цитометрии. J, Процент человеческих клеток AML, привитых к указанным органам, анализировали с помощью проточной цитометрии на 21 день после трансплантации.
Обсуждение
Несмотря на возросшее понимание биологии ОМЛ, методы лечения антителами, непосредственно нацеленные на антигены, экспрессируемые на клетках ОМЛ, с соответствующей специфичностью и функциональностью, еще не изучены.Например, антитело к CD33 (гемтузумаб озогамицин, конъюгат антитело CD33-лекарственное средство) находилось в клинике более 20 лет и показало ограниченный успех в лечении ОМЛ (7, 32). Для лечения ОМЛ срочно необходимы методы лечения, направленные против новых мишеней. Как мы уже сообщали, LILRB4 является поверхностным маркером моноцитарного ОМЛ. Экспрессия LILRB4 в основном наблюдается на нормальных моноцитарных клетках, но повышена на моноцитарных клетках AML. Кроме того, мы показали, что активация LILRB4 внеклеточным связывающим белком APOE ингибирует активацию Т-клеток и поддерживает инфильтрацию лейкозных клеток (17).Вместе эти характеристики делают LILRB4 неотразимой мишенью для разработки терапевтических средств против AML.
mAb были подтверждены как высокоэффективное лекарственное средство для лечения рака. Эти антитела особенно хорошо подходят для блокирования механизмов заболевания, вызванных взаимодействием лиганд-рецептор. В этом исследовании мы стремились создать mAb, блокирующие LILRB4 / APOE, и проверить, обладают ли эти mAb анти-AML активностью in vitro и in vivo . Мы создали комплексную панель кроличьих mAb LILRB4, используя стратегию, оптимизированную в этом исследовании.Эта стратегия включала выделение, культивирование и клонирование В-клеток памяти. Ведущее mAb 128-3 было выбрано на основании его специфичности к LILRB4 и способности блокировать передачу сигналов нижестоящего LILRB4 / APOE. Это ведущее антитело было гуманизировано. Как в линии клеток AML человека, так и в модели мыши NSG первичного ксенотрансплантата клеток AML человека гуманизированное антитело h228-3 проявляло сильную анти-AML активность. Поскольку мыши NSG являются иммунодефицитными и лишены Т-клеток, активность h228-3 мышей NSG против AML, вероятно, является результатом ингибирования миграции раковых клеток и Fc-опосредованных иммунных эффекторных функций, таких как ADCC и ADCP.LILRB4 содержит два Ig-подобных мотива в своих мотивах ECD и ITIM в своем внутриклеточном домене, которые являются отличительными чертами рецепторов, ингибирующих контрольные точки иммунитета (33). Мы стремились определить функцию ингибирования иммунных контрольных точек LILRB4. Для исследований, посвященных этому вопросу, мы выбрали модель мыши C57BL / 6 аллотрансплантата мышиных клеток AML, экспрессирующую человеческий LILRB4. В этой мышиной модели Т-клетки имеют решающее значение для активности против AML. Истощение Т-клеток обработкой антителом CD8 снижает активность h228-3 против AML.Этот результат согласуется с выводами о том, что противоопухолевая эффективность других антител, нацеленных на рецептор, таких как EGFR, CD47, PD-1 и CTLA-4, зависит от Т-клеток. Истощение Т-клеток резко снижает противоопухолевую активность этих терапевтических антител (34–37).
Наши исследования показали, что анти-AML-активность направленного на LILRB4 антитела h228-3 обладает множеством механизмов: (i) обращение супрессии Т-клеток, опосредованной LILRB4; (ii) ингибирование инфильтрации клеточной ткани AML путем блокирования активации LILRB4 с помощью APOE; и (iii) Fc-опосредованные ADCC и ADCP (фиг.6). Fc-опосредованные иммунные функции являются хорошо изученными механизмами действия для многих видов терапии антителами против рака. В некоторых случаях Fc-опосредованные иммунные эффекторные функции избегают, чтобы минимизировать риск истощения нормальных иммунных клеток. Например, большинство одобренных в настоящее время иммуномодулирующих антител, таких как пембролизумаб и ниволумаб, относятся к изотипу IgG4, который имеет низкое связывание или полное отсутствие связывания с рецепторами Fc гамма (FcγR), которые запускают ADCC или ADCP. LILRB4 экспрессируется на моноцитарных клетках AML; нормальные моноциты и дендритные клетки имеют более низкую экспрессию рецептора по сравнению с моноцитарными клетками AML (8).Следовательно, Fc-опосредованные иммунные функции h228-3 могут использоваться для повышения эффективности против AML с потенциально минимальными побочными эффектами из-за зависимости Fc-опосредованных эффекторных функций от числа копий рецептора.
Рисунок 6.Множественные механизмы h228-3 вносят вклад в активность против AML.
Иммуноглобулиноподобные рецепторы лейкоцитов подразделяются на два семейства иммунорегуляторных рецепторов: LILRB (B1-B5) и LILRA (A1-A6). LILRB содержат цитоплазматические хвосты с ITIM, которые дают отрицательные сигналы (8, 38).LILRA имеют короткие цитоплазматические домены, лишенные сигнальных мотивов. LILRA передают активирующие сигналы путем связывания с иммунорецепторными тирозиновыми мотивами активации гамма-цепи FcR (8, 38). LILRB и LILRA экспрессируются на широком спектре типов кроветворных клеток, таких как макрофаги, дендритные клетки, NK-клетки, базофилы и эозинофилы (8, 38). Человеческие гены, кодирующие эти рецепторы, обнаруживаются в кластере генов в хромосомной области 19q13.4. Филогенетический анализ показал, что эти рецепторы обладают высокой степенью сходства последовательностей D1.Поскольку все члены семейства LILR экспрессируются на нормальных иммунных клетках и гематопоэтических клетках, неспецифическое связывание терапевтического антитела LILRB4 может привести к серьезным побочным эффектам. Чтобы получить LILRB4-специфические антитела, мы охарактеризовали большую панель LILRB4-связывающих антител по их аффинности связывания со всеми 11 членами семейства LILR, созданными в виде слитых белков Fc. Высокая специфичность h228-3 к LILRB4 должна повысить профиль безопасности антитела в качестве терапии ОМЛ; это необходимо будет подтвердить в клинических исследованиях.
Таким образом, наше исследование продемонстрировало, что LILRB4 является жизнеспособной лекарственной мишенью для моноцитарного AML. LILRB4-специфичное блокирующее антитело h228-3 продемонстрировало эффективность против AML в моделях in vitro и in vivo . Механистические исследования выявили по крайней мере четыре способа действия анти-AML-эффективности h228-3: включая ингибирование инфильтрации AML-клеток, стимуляцию активации T-клеток, ADCC и ADCP. Взятые вместе, результаты этой работы открывают двери для разработки направленных на LILRB4 антител, таких как h228-3, для лечения моноцитарного AML.
Раскрытие информации о потенциальных конфликтах интересов
X.C. Ляо владеет долей собственности (включая акции, патенты и т. Д.) В Immune-Onc Therapeutics, Inc. Н. Чжан сообщает о получении гранта на коммерческое исследование от Immune-Onc Therapeutics и имеет долю владения патентом на антитела LILRB4 и опционом на акции от Immune. -Onc Therapeutics. С. Чжан сообщает о получении другой коммерческой исследовательской поддержки, поскольку соглашение о спонсируемых исследованиях с Immune-Onc Therapeutics Inc., имеет долю владения патентами, лицензированными для Immune-Onc Therapeutics Inc., и является консультантом / членом консультативного совета компании Immune-Onc Therapeutics Inc. З. Ан имеет долю владения (включая акции, патенты и т. д.) в ImmuneOnc и является членом консультативного совета этой же компании. Другие авторы не сообщали о потенциальных конфликтах интересов.
Вклад авторов
Концепция и дизайн: X. Gui, M. Deng, X.C. Ляо, Н. Чжан, C.C. Zhang, Z. An
Разработка методологии: X. Gui, M. Deng, Y. Xu, T.Хуанг, Н. Ся, Н. Чжан
Сбор данных (предоставленные животные, приобретенные и контролируемые пациенты, предоставленные помещения и т. Д.): X. Gui, M. Deng, H. Song, Y. Chen, Z Li , L. He, F. Huang, Y. Xu, Y. Anami, H. Yu, C. Yu, T. Huang, K. Tsuchikama, N. Zhang
Анализ и интерпретация данных (например, статистический анализ, биостатистика, вычислительный анализ): X. Gui, M. Deng, H. Song, Y. Chen, L. He, Y. Xu, H. Yu, Z. Yuan, Q. Wang, Y. Chai, T. Huang , N. Zhang
Написание, рецензирование и / или исправление рукописи: X.Гуй, М. Дэн, Х. Сун, Т. Хуан, Ю. Ши, X.C. Ляо, Г.Ф. Гао, Н. Чжан, C.C. Zhang, Z. An
Административная, техническая или материальная поддержка (т. Е. Отчетность или систематизация данных, построение баз данных): X. Gui, M. Deng, Y. Chen, J. Xie, L. Li, N. Ся, GF Гао, Н. Чжан, З. Ань
Руководитель исследования: X.C. Ляо, Г.Ф. Гао, Н. Чжан, C.C. Чжан, З. Ань
Благодарности
Мы очень благодарны доктору Сюэцзюнь Фань, доктору Вэй Сюн и госпоже.Хуэй Дэну за техническую помощь и полезные советы. Мы благодарим доктора Джорджину Т. Салазар за редактирование рукописи. Эта работа была поддержана Научно-исследовательским институтом профилактики рака Техаса (RP140402, DP150056, RP180435 и RP150551), Фондом Уэлча (AU-0042-20030616 и I-1834) и Национальным институтом рака (1R01CA172268).
Расходы на публикацию этой статьи были частично покрыты за счет оплаты страницы. Таким образом, данная статья должна быть помечена как реклама в соответствии с 18 U.S.C. Раздел 1734 исключительно для того, чтобы указать на этот факт.
Сноски
Примечание: Дополнительные данные для этой статьи доступны на сайте Cancer Immunology Research Online (http://cancerimmunolres.aacrjournals.org/).
Cancer Immunol Res 2019; 7: 1244–57
- Получено 15 января 2019 г.
- Исправление получено 29 марта 2019 г.
- Принято 12 июня 2019 г.
- Опубликовано впервые 18 июня 2019 г.
- © Американская ассоциация исследований рака, 2019 г.
% PDF-1.3 % 464 0 объект > эндобдж xref 464 73 0000000016 00000 н. 0000002723 00000 н. 0000002825 00000 н. 0000003481 00000 п. 0000004024 00000 н. 0000004379 00000 п. 0000004632 00000 н. 0000004744 00000 н. 0000004858 00000 н. 0000004895 00000 н. 0000004932 00000 н. 0000007251 00000 н. 0000009540 00000 н. 0000009727 00000 н. 0000012020 00000 н. 0000014034 00000 п. 0000016004 00000 п. 0000018123 00000 п. 0000018207 00000 п. 0000018493 00000 п. 0000019169 00000 п. 0000019258 00000 п. 0000019542 00000 п. 0000020158 00000 п. 0000020447 00000 п. 0000020902 00000 н. 0000021182 00000 п. 0000021620 00000 н. 0000021901 00000 п. 0000022430 00000 н. 0000022570 00000 п. 0000022707 00000 п. 0000023351 00000 п. 0000023879 00000 п. 0000024483 00000 п. 0000024985 00000 п. 0000025413 00000 п. 0000025902 00000 п. 0000026188 00000 п. 0000026642 00000 п. 0000029021 00000 н. 0000030712 00000 п. 0000419014 00000 н. 0000419051 00000 н. 0000421587 00000 н. 0000426058 00000 н. 0000442839 00000 н. 0000443194 00000 н. 0000445498 00000 п. 0000448164 00000 н. 0000451530 00000 н. 0000456777 00000 н. 0000459152 00000 п. 0000637519 00000 п. 0000639896 00000 н. 0001467488 00000 п. 0001467981 00000 п. 0001468248 00000 п. 0001468318 00000 п. 0001468474 00000 п. 0001468501 00000 п. 0001468809 00000 п. 0001469209 00000 п. 0001469465 00000 п. 0001469535 00000 п. 0001469680 00000 п. 0001469707 00000 п.