Сн 3057 84: » , — » (. 30.07.1984 N 3057-84) ( » /
Законы :: Воздействие шума на человека (13.140)
ГОСТ 26820-86 Установки силовые вспомогательные пассажирских и транспортных самолетов. Допустимые уровни шума, создаваемого на местности, и метод их определения Постановление Госстандарта СССР от 13.02.1986 N 332 ГОСТ от 13.02.1986 N 26820-86
читать полностью
ГОСТ 12.2.024-87 ССБТ. Шум. Трансформаторы силовые масляные. Нормы и методы контроля Постановление Госстандарта СССР от 23.10.1987 N 4002 ГОСТ от 23.10.1987 N 12.2.024-87
читать полностью
СН 2.2.4/2.1.8.562-96 Шум на рабочих местах, в помещениях жилых, общественных зданий и на территории жилой застройки. Санитарные нормы Постановление Госкомсанэпиднадзора России от 31.10.1996 N 36 Санитарные правила (СП) от 31.10.1996 N 2.2.4/2.1.8.562-96
читать полностью
 2.4.548-96 Гигиенические требования к микроклимату производственных помещений Постановление Госкомсанэпиднадзора России от 01.10.1996 N 21 СанПиН от 01.10.1996 N 2.2.4.548-96
2.4.548-96 Гигиенические требования к микроклимату производственных помещений Постановление Госкомсанэпиднадзора России от 01.10.1996 N 21 СанПиН от 01.10.1996 N 2.2.4.548-96читать полностью
МСанПиН 001-96 Санитарные нормы допустимых уровней физических факторов при применении товаров народного потребления в бытовых условиях. Межгосударственные санитарные правила и нормы Постановление Госкомсанэпиднадзора России от 19.01.1996 N 2 МСанПиН от 19.01.1996 N 001-96
читать полностью
читать полностью
СН 2. 2.4/2.1.8.583-96 Инфразвук на рабочих местах, в жилых и общественных помещениях и на территории жилой застройки Постановление Госкомсанэпиднадзора России от 31.10.1996 N 52 Санитарные правила (СП) от 31.10.1996 N 2.2.4/2.1.8.583-96
2.4/2.1.8.583-96 Инфразвук на рабочих местах, в жилых и общественных помещениях и на территории жилой застройки Постановление Госкомсанэпиднадзора России от 31.10.1996 N 52 Санитарные правила (СП) от 31.10.1996 N 2.2.4/2.1.8.583-96
читать полностью
СН 2.5.2.047-96 Уровни шума на морских судах. Санитарные нормы Постановление Госкомсанэпиднадзора России от 21.02.1996 N 3 Санитарные правила (СП) от 21.02.1996 N 2.5.2.047-96
читать полностью
Методические указания по проведению измерений и гигиенической оценки шумов на рабочих местах Приказ Главного государственного санитарного врача СССР от 25.04.1978 N 1844-78
читать полностью
Методические указания по гигиенической оценке производственной и непроизводственной шумовой нагрузки Приказ Главного государственного санитарного врача СССР от 11.
читать полностью
Методические рекомендации по дозной оценке производственных шумов Приказ Главного государственного санитарного врача СССР от 29.07.1982 N 2908-82
читать полностью
Санитарные нормы допустимого шума, создаваемого изделиями медицинской техники в помещениях лечебно-профилактических учреждений Санитарные правила (СП) от 30.07.1984 N 3057-84 Приказ Главного государственного санитарного врача СССР от 30.07.1984 N 3057-84
читать полностью
Методические рекомендации по борьбе с шумом и вибрацией на предприятиях черной металлургии Приказ Главного государственного санитарного врача СССР от 06.04.1984 N 2986-84
читать полностью
ГОСТ 31296.1-2005 (ИСО 1996-1:2003) Шум. Описание, измерение и оценка шума на местности. Часть 1. Основные величины и процедуры оценки Приказ Ростехрегулирования от 20.07.2006 N 136-ст ГОСТ от 20.07.2006 N 31296.1-2005
Описание, измерение и оценка шума на местности. Часть 1. Основные величины и процедуры оценки Приказ Ростехрегулирования от 20.07.2006 N 136-ст ГОСТ от 20.07.2006 N 31296.1-2005
читать полностью
РД 31.81.81-90 Рекомендации по снижению шума на судах морского флота Письмо Минморфлота СССР от 04.04.1991 N СМ-56/769 РД от 04.04.1991 N 31.81.81-90
читать полностью
Документы ОИ
Разрешительные документы на вид деятельности
Аттестат аккредитации RA.RU.710116 выдан 30 декабря 2015 г.
Перечень нормативно-методических документов, устанавливающих требования к объектам инспекции, методам инспекции
Федеральный закон от 30.03.1999 № 52-ФЗ с изменениями на 3 августа 2018 года «О санитарно-эпидемиологическом благополучии населения»
Федеральный закон от 09. 01.1996 № 3-ФЗ с изменениями на 19 июля 2011 года «О радиационной безопасности населения»
01.1996 № 3-ФЗ с изменениями на 19 июля 2011 года «О радиационной безопасности населения»
Федеральный закон от 04.05.1999 № 96-ФЗ с изменениями на 28 декабря 2017 года «Об охране атмосферного воздуха»
Федеральный закон от 10.01.2002 № 7-ФЗ с изменениями на 31 декабря 2017 года «Об охране окружающей среды»
Приказ Минздравсоцразвития РФ от 12.04.2011 № 302н с изменениями на 6 февраля 2018 года «Об утверждении перечней вредных и (или) опасных производственных факторов и работ, при выполнении которых проводятся обязательные предварительные и периодические медицинские осмотры (обследования), и Порядка проведения обязательных предварительных и периодических медицинских осмотров (обследований) работников, занятых на тяжелых работах и на работах с вредными и (или) опасными условиями труда»
ТР ТС 022/2011 с изменениями на 20 декабря 2017 года «Пищевая продукция в части ее маркировки»
ТР ТС 023/2011 «Технический регламент на соковую продукцию из фруктов и овощей»
ТР ТС 024/2011 с изменениями на 23 апреля 2015 года «Технический регламент на масложировую продукцию»
ТР ТС 027/2012 «О безопасности отдельных видов специализированной пищевой продукции, в том числе диетического лечебного и диетического профилактического питания»
ТР ТС 029/2012 с изменениями на 18 сентября 2014 года «Требования безопасности пищевых добавок, ароматизаторов и технологических вспомогательных средств»
ТР ТС 034/2013 «О безопасности мяса и мясной продукции»
Единые санитарно-эпидемиологические и гигиенические требования к товарам, подлежащим санитарно-эпидемиологическому контролю (надзору) с изменениями на 10 мая 2018 года утв.
 Решением Таможенного союза от 28.05.2010 г. № 299
Решением Таможенного союза от 28.05.2010 г. № 299 СанПиН 2.3.2.1078-01 с изменениями на 6 июля 2011года «Гигиенические требования безопасности и пищевой ценности пищевых продуктов»
МУК 2.6.1.1194-03 «Радиационный контроль. Стронций-90 и цезий-137. Пищевые продукты. Отбор проб, анализ и гигиеническая оценка»
СанПиН 2.2.4.548-96 «Гигиенические требования к микроклимату производственных помещений»
СанПиН 2.2.4/2.1.8.582-96 «Гигиенические требования при работах с источниками воздушного и контактного ультразвука промышленного, медицинского и бытового назначения»
СанПин 2.5.2.703-98 «Суда внутреннего и смешанного (река-море) плавания»
СанПиН 2.6.1.993-00 «Гигиенические требования к обеспечению радиационной безопасности при заготовке и реализации металлолома»
 6.1.993-00 «Гигиенические требования к обеспечению радиационной безопасности при заготовке и реализации металлолома»
6.1.993-00 «Гигиенические требования к обеспечению радиационной безопасности при заготовке и реализации металлолома» СанПиН 2.6.1.3287-15 «Санитарно — эпидемиологические требования к обращению с радиоизотопными приборами и их устройству»
СанПиН 2.1.4.1074 – 01 с изменениями на 28 июня 2010 года «Питьевая вода. Гигиенические требования к качеству питьевой воды централизованных систем питьевого водоснабжения. Контроль качества. Гигиенические требования к обеспечению безопасности систем горячего водоснабжения»
СанПиН 2.1.8/2.2.4.1190-03 «Гигиенические требования к размещению и эксплуатации средств сухопутной подвижной радиосвязи»
СанПиН 2.2.4.3359-16 «Санитарно-эпидемиологические требования к физическим факторам на рабочих местах»
СанПиН 2.6.1.1192-03 с изменениями на 14 февраля 2006 года «Гигиенические требования к устройству и эксплуатации рентгеновских кабинетов, аппаратов и проведению рентгенологических исследований»
СанПиН 2.
 6.1.1281-03 «Санитарные правила по радиационной безопасности персонала и населения при транспортировании радиоактивных материалов (веществ)»
6.1.1281-03 «Санитарные правила по радиационной безопасности персонала и населения при транспортировании радиоактивных материалов (веществ)» СанПиН 2.2.4.1329-03 «Требования по защите персонала от воздействия импульсных электромагнитных полей»
СанПиН 2.2.2.1332-03 с изменениями на 7 сентября 2010 года «Гигиенические требования к организации работы на копировально-множительной технике»
СанПиН 2.2.2/2.4.1340-03 с изменениями на 21 июня 2016 года «Гигиенические требования к персональным электронно-вычислительным машинам и организации работы»
СанПиН 2.1.8/2.2.4.1383-03 с изменениями на 19 декабря 2007 года «Гигиенические требования к размещению и эксплуатации передающих радиотехнических объектов»
СанПиН 2.6.1.2523-09 (НРБ-99/2009) «Нормы радиационной безопасности»
МУ 2.1.674-97 «Санитарно-гигиеническая оценка стройматериалов с добавлением промотходов»
МУ 2.1.2.1829-04 «Санитарно-гигиеническая оценка полимерных и полимерсодержащих строительных материалов и конструкций, предназначенных для применения в строительстве жилых, общественных и промышленных зданий»
СанПиН 2.
 2.4/2.1.8.582-96 «Гигиенические требования при работах с источниками воздушного и контактного ультразвука промышленного, медицинского и бытового назначения»
2.4/2.1.8.582-96 «Гигиенические требования при работах с источниками воздушного и контактного ультразвука промышленного, медицинского и бытового назначения» СанПиН 2.1.3.2630-10 с изменениями на 10 июня 2016 года «Санитарно-эпидемиологические требования к организациям, осуществляющим медицинскую деятельность»
СанПиН 2.1.7.2790-10 «Санитарно-эпидемиологические требования к обращению с медицинскими отходами»
СанПиН 2.1.2.2631-10 с изменениями на 10 июня 2016 года «Санитарно-эпидемиологические требования к размещению, устройству, оборудованию, содержанию и режиму работы организаций коммунально-бытового назначения, оказывающих парикмахерские и косметические услуги»
СанПиН 2.1.2.2645-10 с изменениями от 27 декабря 2010 года «Санитарно-эпидемиологические требования к условиям проживания в жилых зданиях и помещениях»
СП 2.6.1.2612-10 (ОСПОРБ 99/2010) с изменениями на 19 сентября 2013 года «Основные санитарные правила обеспечения радиационной безопасности»
СН 2.
 2.4/2.1.8.562-96 «Шум на рабочих местах, в помещениях жилых, общественных зданий и на территории жилой застройки»
2.4/2.1.8.562-96 «Шум на рабочих местах, в помещениях жилых, общественных зданий и на территории жилой застройки» СН 2.2.4./2.1.8.566-96 «Производственная вибрация, вибрация в помещениях жилых и общественных зданий. Санитарные нормы»
СН 3057-84 «Санитарные нормы допустимого шума, создаваемого изделиями медицинской техники в помещениях лечебно-профилактических учреждений»
ГН 2.2.5.3532-18 «Предельно допустимые концентрации (ПДК) вредных веществ в воздухе рабочей зоны»
ГН 2.1.6.3492-17 «Предельно допустимые концентрации (ПДК) загрязняющих веществ в атмосферном воздухе городских и сельских поселений»
ГН 2.2.5.2308-07 с изменениями на 21 октября 2016 года «Ориентировочные безопасные уровни воздействия (ОБУВ) вредных веществ в воздухе рабочей зоны»
ГН 2.1.6.2309-07 с изменениями на 21 октября 2016 года «Ориентировочные безопасные уровни воздействия (ОБУВ) загрязняющих веществ в атмосферном воздухе населенных мест»
ГН 2.
 1.5.1315-03 с изменениями на 13 июля 2017 года «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
1.5.1315-03 с изменениями на 13 июля 2017 года «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования» ГН 2.1.5.2307-07 с изменениями на 16 сентября 2013 года «Ориентировочные допустимые уровни (ОДУ) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
СанПиН 2.2.2.540-96 «Гигиенические требования к ручным инструментам и организации работ»
СанПиН 2.3.6.1079-01 с изменениями на 10.06.2016г. «Санитарно-эпидемиологические требования к организациям общественного питания, изготовлению и оборотоспособности в них пищевых продуктов и продовольственного сырья»
СанПиН 2.1.4.1110-02 с изменениями от 25 сентября 2014 года «Зоны санитарной охраны источников водоснабжения и водопроводов питьевого назначения»
СанПиН 2.1.4.1175-02 «Гигиенические требования к качеству воды нецентрализованного водоснабжения.
 Санитарная охрана источников»
Санитарная охрана источников» СанПиН 2.1.2.1188-03 «Плавательные бассейны. Гигиенические требования к устройству, эксплуатации и качеству воды. Контроль качества»
СанПиН 2.2.1/2.1.1.1200-03 с изменениями на 25 апреля 2014 года «Санитарно- защитные зоны и санитарная классификация предприятий, сооружений и иных объектов»
СанПиН 2.2.1/2.1.1.1278-03 с изменениями на 15 марта 2010 года «Гигиенические требования к естественному, искусственному и совмещенному освещению жилых и общественных зданий»
СанПиН 2.1.7.1287-03 с изменениями на 25 апреля 2007 года «Санитарно-эпидемиологические требования к качеству почвы»
СанПиН 2.1.7.1322-03 «Гигиенические требования к размещению и обезвреживанию отходов производства и потребления»
СанПиН 2.2.3.1384-03 с изменениями на 3 сентября 2010 года «Гигиенические требования к организации строительного производства и строительных работ»
СанПиН 2.2.3.1385-03 с изменениями на 3 сентября 2010 года «Гигиенические требования к предприятиям производства строительных материалов и конструкций»
СанПиН 2.
 6.1.2800-10 «Гигиенические требования по ограничению облучения населения за счет природных источников ионизирующего излучения»
6.1.2800-10 «Гигиенические требования по ограничению облучения населения за счет природных источников ионизирующего излучения» СанПиН 2.1.5.2582-10 «Санитарно-эпидемиологические требования к охране прибрежных вод морей от загрязнения в местах водопользования населения»
СанПиН 2.4.3.1186-03 с изменениями на 4 марта 2011 года «Санитарно-эпидемиологические требования к организации учебно-производственного процесса в образовательных учреждениях начального профессионального образования»
СанПиН 2.4.5.2409-08 «Санитарно-эпидемиологические требования к организации питания обучающихся в общеобразовательных учреждениях, учреждениях начального и среднего профессионального образования»
СанПиН 2.4.4.2599-10 «Гигиенические требования к устройству, содержанию и организации режима работы в оздоровительных учреждениях с дневным пребыванием детей в период каникул»
СанПиН 2.4.2.2821-10 с изменениями на 24 ноября 2015 года «Санитарно-эпидемиологические требования к условиям и организации обучения в общеобразовательных учреждениях»
СанПиН 2.
 4.1.3049-13 с изменениями на 27 августа 2015 года «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы дошкольных образовательных организаций»
4.1.3049-13 с изменениями на 27 августа 2015 года «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы дошкольных образовательных организаций» СанПиН 2.4.4.3172-14 «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы образовательных организаций дополнительного образования детей»
СанПиН 2.4.4.3155-13 с изменениями на 22 марта 2017 года «Санитарно-эпидемиологические требования к устройству, содержанию и организации работы стационарных организаций отдыха и оздоровления детей
СанПиН 2.4.2.2843-11 «Санитарно-эпидемиологические требования к устройству, содержанию и организации работы детских санаториев»
СП 2.4.3259-15 «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы организаций для детей-сирот и детей, оставшихся без попечения родителей»
СанПиН 2.6.1.3164-14 «Гигиенические требования по обеспечению радиационной безопасности при рентгеновской дефектоскопии»
СанПиН 4690-88 «Санитарные правила содержания территорий населенных мест»
СП 2.
 1.7.1038-01 «Гигиенические требования к устройству и содержанию полигонов для твердых бытовых отходов»
1.7.1038-01 «Гигиенические требования к устройству и содержанию полигонов для твердых бытовых отходов» СП 1.1.1058-01 с изменениями на 27 марта 2007 года «Организация и проведение производственного контроля за соблюдением санитарных правил и выполнением санитарно-противоэпидемических (профилактических) мероприятий»
СП 2.1.5.1059-01 «Гигиенические требования к охране подземных вод от загрязнения»
СП 2.3.6.1066-01 с изменениями на 3 мая 2007 года «Санитарно-эпидемиологические требования к организациям торговли и обороту в них продовольственного сырья и пищевых продуктов»
СП 2.2.2506-09 «Гигиенические требования к организациям химической чистки изделий»
СП 2.2.1.1312-03 с изменениями на 17 мая 2010 года «Гигиенические требования к проектированию вновь строящихся и реконструируемых промышленных предприятий»
СП 2.2.2.1327-03 «Гигиенические требования к организации технологических процессов, производственному оборудованию и рабочему инструменту»
СП 2.
 1.2.2844-11 «Санитарно-эпидемиологические требования к устройству, оборудованию и содержанию общежитий для работников организаций и обучающихся образовательных учреждений»
1.2.2844-11 «Санитарно-эпидемиологические требования к устройству, оборудованию и содержанию общежитий для работников организаций и обучающихся образовательных учреждений» СП 4616-88 «Санитарные правила по гигиене труда водителей автомобилей»
СанПиН 2.3.4.050-96 «Производство и реализация рыбной продукции»
СанПиН 2.2.0.555-96 «Гигиенические требования к условиям труда женщин»
СанПиН 2.1.6.1032-01 «Гигиенические требования к обеспечению качества атмосферного воздуха населенных мест»
СанПиН 2.1.4.1175-02 «Гигиенические требования к качеству воды нецентрализованного водоснабжения. Санитарная охрана источников»
СП 3.5.1378-03 «Санитарно-эпидемиологические требования к организации и осуществлению дезинфекционной деятельности»
СП 3244-85 «Санитарные правила для предприятий пивоваренной и безалкогольной промышленности»
ГН 2.1.7.2041-06 с изменениями на 26 июня 2017 года «Предельно допустимые концентрации (ПДК) химических веществ в почве»
ГН 2.
 1.7.2511-09 «Ориентировочно допустимые концентрации (ОДК) химических веществ в почве»
1.7.2511-09 «Ориентировочно допустимые концентрации (ОДК) химических веществ в почве» ГН 2.1.5.1315-03 с изменениями на 13 июля 2017 года «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
ГН 2.1.5.2307-07 с изменениями на 16 сентября 2013года «Ориентировочные допустимые уровни (ОДУ) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
МУ 2.6.1.1981-05 «Радиационный контроль и гигиеническая оценка источников питьевого водоснабжения и питьевой воды по показателям радиационной безопасности. Оптимизация защитных мероприятий источников питьевого водоснабжения с повышенным содержанием радионуклидов»
МУ 2.6.1.2713-10 «Радиационный контроль и гигиеническая оценка источников питьевого водоснабжения и питьевой воды по показателям радиационной безопасности. Оптимизация защитных мероприятий источников питьевого водоснабжения с повышенным содержанием радионуклидов.
 Изменение № 1 к МУ 2.6.1.1981-05»
Изменение № 1 к МУ 2.6.1.1981-05» МУК 4.3.2756-10 «Методические указания по измерению и оценке микроклимата производственных помещений»
МУК 4.3.2194-07 «Контроль уровня шума на территории жилой застройки, в жилых и общественных зданиях и помещениях»
МУК 4.3.2491-09 «Гигиеническая оценка электрических и магнитных полей промышленной частоты (50 Гц) в производственных условиях»
МУ 2.2.4.706-98/МУ ОТ РМ 01-98 «Оценка освещения рабочих мест»
МУ 2.1.674-97 «Санитарно-гигиеническая оценка стройматериалов с добавлением промотходов»
МУ 2.1.2.1829-04 «Санитарно-гигиеническая оценка полимерных и полимерсодержащих строительных материалов и конструкций, предназначенных для применения в строительстве жилых, общественных и промышленных зданий»
МУ 2.2.8/2.6.1.67-02 «Организация вентиляции на радиационных объектах»
МУ 4425-87 «Санитарно-гигиенический контроль систем вентиляции производственных помещений.
 Методические указания»
Методические указания» ГОСТ Р ИСО 9612-2013 «Акустика. Измерения шума для оценки его воздействия на человека. Метод измерений на рабочих местах»
МУ 4.3.2320-08 «Порядок подготовки и оформления санитарно-эпидемиологических заключений на передающие радиотехнические объекты»
Р 2.2.2006-05 «Гигиена труда. Руководство по гигиенической оценке факторов рабочей среды и трудового процесса. Критерии и классификация условий труда»
МУ 1844-78 «Методические указания по проведению измерений и гигиенической оценки шумов на рабочих местах»
МУ 4435-87 «Методические указания по гигиенической оценке производственной и непроизводственной шумовой нагрузки»
ГОСТ 31191.1-2004 «Вибрация и удар. Измерение общей вибрации и оценка ее воздействия на человека. Часть 1. Общие требования»
ГОСТ 31191.2-2004 (ИСО 2631-2:2003) «Вибрация и удар. Измерение общей вибрации и оценка ее воздействия на человека. Часть 2. Вибрация внутри зданий»
ГОСТ Р 54944-2012 «Здания и сооружения.
 Методы измерения освещенности»
Методы измерения освещенности» ГН 2.1.8/2.2.4.2262-07 «Предельно допустимые уровни магнитных полей частотой 50 Гц в помещениях жилых, общественных зданий и на селитебных территориях»
МУ 2.6.1.2838-11 «Радиационный контроль и санитарно-эпидемиологическая оценка жилых, общественных и производственных зданий и сооружений после окончания их строительства, капитального ремонта, реконструкции по показателям радиационной безопасности»
СанПиН 2.6.1.2800-10 «Гигиенические требования по ограничению облучения населения за счет источников ионизирующего излучения»
ГОСТ 12.4.077-79 «Система стандартов безопасности труда (ССБТ). Ультразвук. Метод измерения звукового давления на рабочих местах»
МУ 4.3.1517-03 «Санитарно-эпидемиологическая оценка и эксплуатация аэроионизирующего оборудования»
СанПиН 2.2.4.1294-03 «Гигиенические требования к аэроионному составу воздуха производственных и общественных помещений»
МУ 4109-86 «Методические указания по определению электромагнитного поля воздушных высоковольтных линий электропередачи и гигиенические требования к их размещению»
МУ 2375-81 «Методические указания по предупредительному санитарному надзору при проектировании, строительстве, переоборудовании и приемке в эксплуатацию судов морского, смешанного и внутреннего плавания СССР и других объектов и сооружений, приравненных к ним»
МУ 1939-78 «Методические указания по проведению санитарно-химических исследований воздушной среды судовых помещений, оборудованных полимерными материалами»
МУ 3169-84 «Организация и проведение санитарно-гигиенического надзора при перевозке и перегрузке пылящих навалочных грузов в портах»
МУ 4260-87 «Методические указания по осуществлению государственного санитарного надзора за судовыми установками очистки и обеззараживания сточных вод»
МУ 2639-82 «Методические указания по организации и проведению текущего санитарного надзора за эксплуатируемыми судами и другими плавучими средствами»
МУ 2.
 1.2.694-98 «Использование ультрафиолетового излучения при обеззараживании воды плавательных бассейнов»
1.2.694-98 «Использование ультрафиолетового излучения при обеззараживании воды плавательных бассейнов» МУ 2.3.975-00 «Применение ультрафиолетового бактерицидного излучения для обеззараживания воздушной среды помещений организаций пищевой промышленности, общественного питания и торговли продовольственными товарами»
МУ 2.6.1.1982-05 «Проведение радиационного контроля в рентгеновских кабинетах»
МУ 2.6.1.2398-08 «Радиационный контроль и санитарно-эпидемиологическая оценка земельных участков под строительство жилых домов, зданий и сооружений общественного и производственного назначения в части обеспечения радиационной безопасности»
МУ 2.6.1.2944-11 «Контроль эффективных доз облучения пациентов при проведении медицинских рентгенологических исследований»
МУ 2.6.1.3015-12 «Организация и проведение индивидуального дозиметрического контроля. Персонал медицинских организаций»
МУ 3911-85 «По проведению измерений и гигиенической оценки производственных вибраций»
МУК 2.
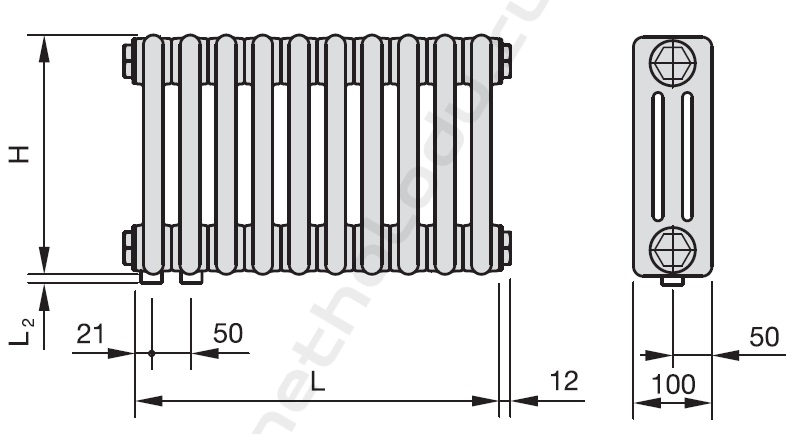 6.1.1087-02 с дополнением от 4 декабря 2006 года «Радиационный контроль металлолома»
6.1.1087-02 с дополнением от 4 декабря 2006 года «Радиационный контроль металлолома» МУК 4.3.1677-03 «Определение уровней электромагнитного поля, создаваемого излучающими техническими средствами телевидения, ЧМ радиовещания и базовых станций сухопутной подвижной радиосвязи»
МУК 4.3.2812-10 «Инструментальный контроль и оценка освещения рабочих мест»
МР 2946-83 «Методические рекомендации по измерению импульсной локальной вибрации»
Р 3.5.1904-04 «Использование ультрафиолетового бактерицидного излучения для обеззараживания воздуха в помещениях»
ГОСТ 2761-84 «Источники централизованного хозяйственно-питьевого водоснабжения. Гигиенические, технические требования и правила выбора»
ГОСТ 16371-2014 «Мебель. Общие технические условия»
ГОСТ 22046-2016 «Мебель для учебных заведений. Общие технические условия»
ГОСТ 26756-2016 «Мебель для предприятий торговли. Общие технические условия»
СанПиН 2.
 1.2882-11 «Гигиенические требования к размещению, устройству и содержанию кладбищ, зданий и сооружений похоронного назначения»
1.2882-11 «Гигиенические требования к размещению, устройству и содержанию кладбищ, зданий и сооружений похоронного назначения» СанПиН 2.1.2.3150-13 «Санитарно-эпидемиологические требования к размещению, устройству, оборудованию, содержанию и режиму работы бань и саун»
СП 1.3.2322-08 с изменениями на 29 июня 2011года «Безопасность работы с микроорганизмами III — IV групп патогенности (опасности) и возбудителями паразитарных инфекций»
СП 3.5.3.3223-14 «Санитарно-эпидемиологические требования к организации и проведению дератизационных мероприятий»
Перечень документов, используемых при выполнении органом инспекции работ по оценке соответствия
Федеральный закон от 30.03.1999 N 52-ФЗ «О санитарно-эпидемиологическом благополучии населения»
Федеральный закон от 22.12.2008 N 268-ФЗ «Технический регламент на табачную продукцию»
Федеральный закон от 1 июля 2017 г. № 135-ФЗ “О внесении изменений в отдельные законодательные акты Российской Федерации в части совершенствования порядка установления и использования приаэродромной территории и санитарно-защитной зоны”
№ 135-ФЗ “О внесении изменений в отдельные законодательные акты Российской Федерации в части совершенствования порядка установления и использования приаэродромной территории и санитарно-защитной зоны”
Федеральный закон от 30.12.2009 N 384-ФЗ «Технический регламент о безопасности зданий и сооружений»
Постановление Правительства РФ от 3 марта 2018 г. № 222 «Об утверждении Правил установления санитарно-защитных зон и использования земельных участков, расположенных в границах санитарно-защитных зон»
ТР ТС 004/2011 «Технический регламент Таможенного Союза. О безопасности низковольтного оборудования»
ТР ТС 005/2011 «Технический регламент Таможенного союза. О безопасности упаковки»
ТР ТС 007/2011 «Технический регламент Таможенного Союза. О безопасности продукции, предназначенной для детей и подростков»
ТР ТС 008/2011 «Технический регламент ТС. О безопасности игрушек»
ТР ТС 009/2011 «Технический регламент Таможенного союза. О безопасности парфюмерно-косметической продукции»
О безопасности парфюмерно-косметической продукции»
ТР ТС 010/2011 «Технический регламент Таможенного союза. О безопасности машин и оборудования»
ТР ТС 015/2011 «Технический регламент Таможенного союза. О безопасности зерна»
ТР ТС 017/2011 «Технический регламент Таможенного союза. О безопасности продукции легкой промышленности»
ТР ТС 019/2011 «Технический регламент Таможенного союза. О безопасности средств индивидуальной защиты»
ТР ТС 021/2011 «Технический регламент Таможенного союза. О безопасности пищевой продукции»
ТР ТС 022/2011 «Технический регламент Таможенного союза. Пищевая продукция в части ее маркировки»
ТР ТС 023/2011 «Технический регламент Таможенного союза. Технический регламент на соковую продукцию из фруктов и овощей»
ТР ТС 024/2011 «Технический регламент Таможенного союза. Технический регламент на масложировую продукцию»
ТР ТС 027/2012 «Технический регламент Таможенного союза. О безопасности отдельных видов специализированной пищевой продукции, в том числе диетического лечебного и диетического профилактического питания»
О безопасности отдельных видов специализированной пищевой продукции, в том числе диетического лечебного и диетического профилактического питания»
ТР ТС 029/2012 «Технический регламент Таможенного союза. Требования безопасности пищевых добавок, ароматизаторов и технологических вспомогательных средств»
ТР ТС 033/2013 «Технический регламент Таможенного союза. О безопасности молока и молочной продукции»
ТР ТС 034/2013 «Технический регламент Таможенного союза. О безопасности мяса и мясной продукции»
Технический регламент Евразийского экономического союза «О безопасности рыбы и рыбной продукции» (ТР ЕАЭС 040/2016), принят Решением Совета Евразийской комиссии от 18.10.2016 N 162
ПРИКАЗ от 19 июля 2007 года N 224 «О санитарно-эпидемиологических экспертизах, обследованиях, исследованиях, испытаниях и токсикологических, гигиенических и иных видах оценок»
СанПиН 2.2.4.3359-16 «Санитарно-эпидемиологические требования к физическим факторам на рабочих местах»
СанПиН 2. 2.4/2.1.8.582-96 «Гигиенические требования при работах с источниками воздушного и контактного ультразвука промышленного, медицинского и бытового назначения»
2.4/2.1.8.582-96 «Гигиенические требования при работах с источниками воздушного и контактного ультразвука промышленного, медицинского и бытового назначения»
СанПиН 2.1.7.573-96 «Гигиенические требования к использованию сточных вод и их осадков для орошения и удобрения»
СанПиН 2.1.8/2.2.4.1190-03 «Гигиенические требования к размещению и эксплуатации средств сухопутной подвижной радиосвязи»
СанПиН 2.2.1/2.1.1.1200-03 «Санитарно-защитные зоны и санитарная классификация предприятий, сооружений и иных объектов»
СанПиН 2.2.1./2.1.1.-2361-08 «Санитарно-защитные зоны и санитарная классификация предприятий, сооружений и иных объектов. Изменение N 1 к СанПиН 2.2.1./2.1.1.1200-03″
СанПиН 2.2.1/2.1.1.2555-09 «Изменение N 2 к СанПиН 2.2.1/2.1.1.1200-03 «Санитарно-защитные зоны и санитарная классификация предприятий, сооружений и иных объектов. Новая редакция»
СанПиН 2.2.1/2.1. 1.2739-10 «Изменения и дополнения N 3 к СанПиН 2.2.1/2.1.1.1200-03 «Санитарно-защитные зоны и санитарная классификация предприятий, сооружений и иных объектов. Новая редакция»
1.2739-10 «Изменения и дополнения N 3 к СанПиН 2.2.1/2.1.1.1200-03 «Санитарно-защитные зоны и санитарная классификация предприятий, сооружений и иных объектов. Новая редакция»
СанПиН 2.1.6.1032-01 «Гигиенические требования к обеспечению качества атмосферного воздуха населенных мест»
СанПиН 1.2.2834-11 «Дополнения и изменения N 1 к СанПиН 1.2.2353-08 «Канцерогенные факторы и основные требования к профилактике канцерогенной опасности»
СанПиН 1.2.2353-08 «Канцерогенные факторы и основные требования к профилактике канцерогенной опасности»
СанПиН 2.2.2/2.4.1340-03 «Гигиенические требования к персональным электронно-вычислительным машинам и организации работы»
СанПиН 2.2.2/2.4.2198-07 «Изменение N 1 к санитарно-эпидемиологическим правилам и нормативам «Гигиенические требования к персональным электронно-вычислительным машинам и организации работы. СанПиН 2.2.2/2.4.1340-03″
СанПиН 2. 2.2.1332 -03 «Гигиенические требования к организации работы на копировально-множительной технике»
2.2.1332 -03 «Гигиенические требования к организации работы на копировально-множительной технике»
СанПиН 2.2.3.2887-11 «Гигиенические требования при производстве и использовании хризотила и хризотилсодержащих материалов»
СанПиН 2.2.2.540-96 «Гигиенические требования к ручным инструментам и организации работ»
СанПиН 2.2.4.1329-03 «Требования по защите персонала от воздействия импульсных электромагнитных полей»
СанПиН 2.2.2.2731-10 «Изменение N 1 к СанПиН 2.2.2.1332-03 «Гигиенические требования к организации работы на копировально-множительной технике»
СанПиН 2.5.1.2423-08 «Гигиенические требования к условиям труда и отдыха для летного состава гражданской авиации»
СанПиН 2.2.2948-11 «Гигиенические требования к организациям, осуществляющим деятельность по добыче и переработке угля (горючих сланцев) и организации работ»
СанПиН 2.2.3.1384-03 «Гигиенические требования к организации строительного производства и строительных работ»
СанПиН 2. 2.3.2733-10 «Изменение N 1 к СанПиН 2.2.3.1384-03 «Гигиенические требования к организации строительного производства и строительных работ»
2.3.2733-10 «Изменение N 1 к СанПиН 2.2.3.1384-03 «Гигиенические требования к организации строительного производства и строительных работ»
СанПиН 2.3.4.050-96 «Производство и реализация рыбной продукции»
СанПиН 2.5.2/2.2.4.1989-06 «Электромагнитные поля на плавательных средствах и морских сооружениях. Гигиенические требования безопасности»
СанПиН 2.6.1.993-00 «Гигиенические требования к обеспечению радиационной безопасности при заготовке и реализации металлолома»
СанПиН 2.6.1.1202-03 «Гигиенические требования к использованию закрытых адионуклидных источников ионизирующего излучения при геофизических работах на буровых скважинах»
СанПиН 2.2.3.1385-03 «Гигиенические требования к предприятиям производства строительных материалов и конструкций»
СанПиН 2.5.2.703-98 «Суда внутреннего и смешенного (река-море) плавания. Раздел 2.1.4. Помещения пищеблока для экипажа. Раздел 3. 2. Организация питания экипажа и пассажиров»
2. Организация питания экипажа и пассажиров»
СанПиН 2.4.6.2553-09 «Санитарно-эпидемиологические требования к безопасности условий труда работников, не достигших 18-летнего возраста»
СанПиН 2.4.3.1186-03 «Санитарно-эпидемиологические требования к организации учебно-производственного процесса в образовательных учреждениях начального профессионального образования»
СанПиН 2.6.1.3289-15 «Гигиенические требования по обеспечению радиационной езопасности при обращении с источниками, генерирующими рентгеновское излучение при ускоряющем напряжении до 150 кВ»
СанПиН 2.1.2.2631-10 «Санитарно-эпидемиологические требования к размещению, устройству, оборудованию, содержанию и режиму работы организаций коммунально-бытового назначения, оказывающих парикмахерские и косметические услуги»
СанПиН 2.4.1.3049-13 «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы дошкольных образовательных организаций»
СанПиН 2. 6.1.1192-03 «Гигиенические требования к устройству и эксплуатации рентгеновских кабинетов, аппаратов и проведению рентгенологических исследований»
6.1.1192-03 «Гигиенические требования к устройству и эксплуатации рентгеновских кабинетов, аппаратов и проведению рентгенологических исследований»
СанПиН 5804-91 «Санитарные нормы и правила устройства и эксплуатации лазеров»
СанПиН 2.2.4.548-96 «Гигиенические требования к микроклимату производственных помещений»
СанПиН 2.1.8/2.2.4.2302-07 «Гигиенические требования к размещению и эксплуатации передающих радиотехнических объектов. Изменение N 1 к СанПиН 2.1.8/2.2.4.1383-03″
СанПиН 2.1.8/2.2.4.1383-03 «Гигиенические требования к размещению и эксплуатации передающих радиотехнических объектов»
СанПиН 2.1.2.2645-10 «Санитарно-эпидемиологические требования к условиям проживания в жилых зданиях и помещениях»
СанПиН 2.2.1/2.1.1.1278-03 «Гигиенические требования к естественному, искусственному и совмещенному освещению жилых и общественных зданий»
СанПиН 2. 2.1/2.1.1.2585-10 «Изменения и дополнения N 1 к санитарным правилам и нормам СанПиН 2.2.1/2.1.1.1278-03 Гигиенические требования к естественному, искусственному и совмещенному освещению жилых и общественных зданий»
2.1/2.1.1.2585-10 «Изменения и дополнения N 1 к санитарным правилам и нормам СанПиН 2.2.1/2.1.1.1278-03 Гигиенические требования к естественному, искусственному и совмещенному освещению жилых и общественных зданий»
СанПиН 2.2.0.555-96 «Гигиенические требования к условиям труда женщин»
СанПиН 2.2.2/2.4.2620-10 «Изменения N 2 к СанПиН 2.2.2/2.4.1340-03 «Гигиенические требования к персональным электронно-вычислительным машинам и организации работы»
СанПиН 2.6.1.3239-14 «Производство и применение радиолюминесцентных источников света с газообразным тритием и изделий на их основе»
СанПиН 2.6.1.3288-15 «Гигиенические требования по обеспечению радиационной безопасности при подготовке и проведении позитронной эмиссионной томографии»
СанПиН 2.6.1.3488-17 «Гигиенические требования по обеспечению радиационной безопасности при обращении с лучевыми досмотровыми установками»
СанПиН 2. 4.2.2821-10 «Санитарно-эпидемиологические требования к условиям и организации обучения в общеобразовательных учреждениях»
4.2.2821-10 «Санитарно-эпидемиологические требования к условиям и организации обучения в общеобразовательных учреждениях»
СанПиН 2.1.4.1074-01 «Питьевая вода. Гигиенические требования к качеству воды централизованных систем питьевого водоснабжения. Контроль качества. Гигиенические требования к обеспечению безопасности систем горячего водоснабжения»
СанПиН 2.1.2.2801-10 «Изменения и дополнения N 1 к СанПиН 2.1.2.2645-10 «Санитарно-эпидемиологические требования к условиям проживания в жилых зданиях и помещениях»
СанПиН 2.1.7.1322-03 «Гигиенические требования к размещению и обезвреживанию отходов производства и потребления»
СанПиН 2.2.1/2.1.1.1076-01 «Гигиенические требования к инсоляции и солнцезащите помещений жилых и общественных зданий и территорий»
СанПиН 2.1.3.2630-10 «Санитарно-эпидемиологические требования к организациям, осуществляющим медицинскую деятельность»
СанПиН 42-128-4690-88 «Санитарные правила содержания территорий населенных мест»
СанПиН 2. 2.2506-09 «Гигиенические требования к организациям химической чистки изделий»
2.2506-09 «Гигиенические требования к организациям химической чистки изделий»
СанПиН 2.2.3.2734-10 «Изменение N 1 к СанПиН 2.2.3.1385-03 «Гигиенические требования к предприятиям производства строительных материалов и конструкций»
СанПиН 2.3.4.551-96 «Производство молока и молочных продуктов»
СанПиН 2.1.2.2646-10 «Санитарно-эпидемиологические требования к устройству, оборудованию, содержанию и режиму работы прачечных»
СанПиН 2.1.2.1188-03 «Плавательные бассейны. Гигиенические требования к устройству, эксплуатации и качеству воды. Контроль качества»
СанПиН 2.1.2.1331-03 «Гигиенические требования к устройству, эксплуатации и качеству воды аквапарков»
СанПиН 2.1.2/3041-96 «Устройство, оборудование и содержание центров временного размещения иммигрантов — иностранных граждан, лиц без гражданства и беженцев»
СанПиН 2.1.2.3150-13 «Санитарно-эпидемиологические требования к размещению, устройству, оборудованию, содержанию и режиму работы бань и саун»
СанПиН 2. 1.7.2790-10 «Санитарно-эпидемиологические требования к обращению с медицинскими отходами»
1.7.2790-10 «Санитарно-эпидемиологические требования к обращению с медицинскими отходами»
СанПиН 2.1.2882-11 «Гигиенические требования к размещению, устройству и содержанию кладбищ, зданий и сооружений похоронного назначения»
СанПиН 2.6.1.2523-09 «Нормы радиационной безопасности НРБ-99/2009″
СанПиН 2.6.1.2800-10 «Гигиенические требования по ограничению облучения населения за счет природных источников ионизирующего излучения»
СанПиН 2.6.1.3287-15 «Санитарно-эпидемиологические требования к обращению с радиоизотопными приборами и их устройству»
СанПиН 2.6.1.3164-14 «Гигиенические требования по обеспечению радиационной безопасности при рентгеновской дефектоскопии»
СанПиН 2.6.1.2368-08 «Гигиенические требования по обеспечению радиационной езопасности при проведении лучевой терапии с помощью открытых радионуклидных источников»
СанПиН 2.6.1.2573-10 «Гигиенические требования к размещению и эксплуатации ускорителей электронов с энергией до 100 МэВ»
СанПиН 2. 6.1.2748-10 «Гигиенические требования по обеспечению радиационной безопасности при работе с источниками неиспользуемого рентгеновского излучения»
6.1.2748-10 «Гигиенические требования по обеспечению радиационной безопасности при работе с источниками неиспользуемого рентгеновского излучения»
СанПиН 2.6.1.2891-11 «Требования радиационной безопасности при производстве, эксплуатации и выводе из эксплуатации (утилизации) медицинской техники, содержащей источники ионизирующего излучения»
СанПиН 2.6.1.3106-13 «Гигиенические требования по обеспечению радиационной безопасности при использовании рентгеновских сканеров для персонального досмотра людей»
СанПиН 2.1.4.1110-02 «Зоны санитарной охраны источников водоснабжения и водопроводов питьевого назначения»
СанПиН 2.1.4.2496-09 «Гигиенические требования к обеспечению безопасности систем горячего водоснабжения. Изменение к СанПиН 2.1.4.1074-01″
СанПиН 2.4.4.3172-14 «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы образовательных организаций дополнительного образования детей»
СанПиН 1. 2.2584-10 «Гигиенические требования к безопасности процессов испытаний, хранения, перевозки, реализации, применения, обезвреживания и утилизации пестицидов и агрохимикатов»
2.2584-10 «Гигиенические требования к безопасности процессов испытаний, хранения, перевозки, реализации, применения, обезвреживания и утилизации пестицидов и агрохимикатов»
СанПиН 3.5.2.3472-17 «Санитарно-эпидемиологические требования к организации и проведению дезинсекционных мероприятий в борьбе с членистоногими, имеющими эпидемиологическое и санитарно-гигиеническое значение»
СанПиН 1.2.1330-03 «Гигиенические требования к производству пестицидов и агрохимикатов»
СанПиН 2.4.3259-15 «Санитарно-эпидемиологические требования к устройству, содержанию и организации режима работы организаций для детей-сирот и детей, оставшихся без попечения родителей»
СанПиН 2.4.1.3147-13 «Санитарно-эпидемиологические требования к дошкольным группам, размещенным в жилых помещениях жилищного фонда»
СанПиН 2.4.2.2843-11 «Санитарно-эпидемиологические требования к устройству, содержанию и организации работы детских санаториев»
СанПиН 2. 1.7.1287-03 «Санитарно-эпидемиологические требования к качеству почвы»
1.7.1287-03 «Санитарно-эпидемиологические требования к качеству почвы»
СанПиН 2.1.7.2197-07 «Изменение N 1 к санитарно-эпидемиологическим правилам и нормативам «Санитарно-эпидемиологические требования к качеству почвы. СанПиН 2.1.7.1287-03″
СанПиН 2.1.5.980-00 «Гигиенические требования к охране поверхностных вод»
СанПиН 2.4.2.3286-15 «Санитарно-эпидемиологические требования к условиям и организации обучения и воспитания в организациях, осуществляющих образовательную деятельность по адаптированным основным общеобразовательным программам для обучающихся с ограниченными возможностями здоровья»
СанПиН 2.4.7.1166-02 «Гигиенические требования к изданиям учебным для общего и начального профессионального образования»
СанПиН 2.4.7.960-00 «Гигиена детей и подростков. Гигиенические требования к изданиям книжным и журнальным для детей и подростков»
СанПиН 2.6.1.2802-10 «Гигиенические требования по обеспечению радиационной безопасности при проведении работ со скважинными генераторами нейтронов»
СанПиН 2. 6.1.2749-10 «Гигиенические требования по обеспечению радиационной безопасности при обращении с радиоизотопными термоэлектрическими генераторами»
6.1.2749-10 «Гигиенические требования по обеспечению радиационной безопасности при обращении с радиоизотопными термоэлектрическими генераторами»
СанПиН 2.4.7/1.1.1286-03 «Гигиенические требования к одежде для детей, подростков и взрослых»
СанПиН 2.4.7/1.1.2651-10 «Дополнения и изменения N 1 к санитарно-эпидемиологическим правилам и нормативам СанПиН 2.4.7/1.1.1286-03 «Гигиенические требования к одежде для детей, подростков и взрослых»
СанПиН 2.1.8/2.2.4.2489-09 «Гипогеомагнитные поля в производственных, жилых и общественных зданиях и сооружениях»
СанПиН 2.6.1.1281-03 «Санитарные правила по радиационной безопасности персонала и населения при транспортировании радиоактивных материалов (веществ)»
СанПиН 2.4.2.2842-11 «Санитарно-эпидемиологические требования к устройству, содержанию и организации работы лагерей труда и отдыха для подростков»
СанПиН 2.4.4.3155-13 «Санитарно-эпидемиологические требования к устройству, содержанию и организации работы стационарных организаций отдыха и оздоровления детей»
СанПиН 2. 4.4.2599-10 «Гигиенические требования к устройству, содержанию и организации режима работы в оздоровительных учреждениях с дневным пребыванием детей в период каникул»
4.4.2599-10 «Гигиенические требования к устройству, содержанию и организации режима работы в оздоровительных учреждениях с дневным пребыванием детей в период каникул»
СанПиН 2.4.4.3048-13 «Санитарно-эпидемиологические требования к устройству и организации работы детских лагерей палаточного типа»
СанПиН 2.2.2/2.4.2732-10 «Изменения N 3 к СанПиН 2.2.2/2.4.1340-03 «Гигиенические требования к персональным электронно-вычислительным машинам и организации работы»
СанПиН 2.1.2.2564-09 «Гигиенические требования к размещению, устройству, оборудованию, содержанию объектов организаций здравоохранения и социального обслуживания, предназначенных для постоянного проживания престарелых и инвалидов, санитарно-гигиеническому и противоэпидемическому режиму их работы»
СанПиН 001-96 «Санитарные нормы допустимых уровней физических факторов при применении товаров народного потребления в бытовых условиях. Межгосударственные санитарные правила и нормы»
СанПиН 2. 3.2.1324-03 «Гигиенические требования к срокам годности и условиям хранения пищевых продуктов»
3.2.1324-03 «Гигиенические требования к срокам годности и условиям хранения пищевых продуктов»
СанПиН 2.3.4.704-98 «Производство спирта этилового ректификованного и ликероводочных изделий»
СанПиН № 1197-74 «Санитарные правила для предприятий по производству растительных масел»
СанПиН 2.3.2.1290-03 «Гигиенические требования к организации производства и оборота биологически активных добавок к пище (БАД)»
СанПиН 42-123-4717-88 «Санитарно-гигиенические нормы «Рекомендуемые (регламентируемые) уровни содержания витаминов в витаминизированных пищевых продуктах»
СанПиН 2.1.4.2652-10 «Изменение N 3 в СанПиН 2.1.4.1074-01 «Питьевая вода. Гигиенические требования к качеству воды централизованных систем питьевого водоснабжения. Контроль качества. Гигиенические требования к обеспечению безопасности систем горячего водоснабжения»
СанПиН 2.1.4.2653-10 «Изменения N 2 к СанПиН 2. 1.4.1116-02 «Питьевая вода. Гигиенические требования к качеству воды, расфасованной в емкости. Контроль качества»
1.4.1116-02 «Питьевая вода. Гигиенические требования к качеству воды, расфасованной в емкости. Контроль качества»
СанПиН 2.1.4.2581-10 «Питьевая вода. Гигиенические требования к качеству воды, расфасованной в емкости. Контроль качества. Изменения N 1 к СанПиН 2.1.4.1116-02″
СанПиН 2.1.4.1116-02 «Питьевая вода. Гигиенические требования к качеству воды, расфасованной в емкости. Контроль качества»
СанПиН 2.1.4.1175-02 «Гигиенические требования к качеству воды нецентрализованного водоснабжения. Санитарная охрана источников»
СанПиН 2.1.4.2580-10 «Изменения N 2 к СанПиН 2.1.4.1074-01 «Питьевая вода. Гигиенические требования к качеству воды централизованных систем питьевого водоснабжения. Контроль качества»
СанПиН 2.3.2.1078-01 «Гигиенические требования безопасности и пищевой ценности пищевых продуктов»
СанПиН 2.3.2.1293-03 «Гигиенические требования по применению пищевых добавок»
СанПиН 42-123-4689-88 «Рекомендуемый состав, критерии и показатели качества заменителей женского молока»
СанПиН 42-123-4717-88 Санитарно-гигиенические нормы «Рекомендуемые (регламентируемые) уровни содержания витаминов в витаминизированных пищевых продуктах»
СП 2. 2.2.1327-03 «Гигиенические требования к организации технологических процессов, производственному оборудованию и рабочему инструменту»
2.2.1327-03 «Гигиенические требования к организации технологических процессов, производственному оборудованию и рабочему инструменту»
СП 2.2.1.1312-03 «Гигиенические требования к проектированию вновь строящихся и реконструируемых промышленных предприятий»
СП № 2550-82 «Предельно допустимые уровни напряженности электромагнитного поля, создаваемого индукционными бытовыми печами, работающими на частоте 20-22 кГц»
СП № 2524-82 «Санитарные правила по сбору, хранению, транспортировке и первичной обработке вторичного сырья»
СП 1.2.1170-02 «Гигиенические требования к безопасности агрохимикатов»
СП 1.1.1058-01 «Организация и проведение производственного контроля за соблюдением санитарных правил и выполнением санитарно-противоэпидемических (профилактических) мероприятий»
СП № 952-72 «Санитарные правила организации процессов пайки мелких изделий сплавами, содержащими свинец»
СП № 991-72 «Санитарные правила при окрасочных работах с применением ручных распылителей. Санитарно-гигиеническая характеристика условий труда»
Санитарно-гигиеническая характеристика условий труда»
СП № 1009-73 «Санитарные правила при сварке, наплавке и резке металлов»
СП № 3935-85 «Санитарные правила при работе со смазочно-охлаждающими жидкостями и технологическими смазками»
СП № 5159-89 «Санитарные правила при производстве и применении эпоксидных смол и материалов на их основе»
СП № 5160-89 «Санитарные правила для механических цехов (обработка металлов резанием) «
СП № 4224-86 «Санитарные правила для процессов обработки металлов резанием»
СП № 2400-81 «Санитарные правила по устройству, оборудованию и эксплуатации предприятий производства стекловолокна и стеклопластиков»
СП № 1451-76 «Санитарные правила для предприятий по производству сварочных материалов (электродов, порошковой проволоки и флюсов) «
СП № 1983-79 «Санитарные правила для производств основных свинецсодержащих пигментов»
СП № 2528-82 «Санитарные правила для предприятий цветной металлургии»
СП № 4079-86 «Санитарные правила для предприятий по производству лекарственных препаратов»
СП № 4155-86 «Санитарные правила для производства фосфора и его неорганических соединений»
СП № 4156-86 «Санитарные правила для нефтяной промышленности»
СП № 4616-88 «Санитарные правила по гигиене труда водителей автомобилей»
СП № 4783-88 «Санитарные правила для производств синтетических полимерных материалов и предприятий по их переработке»
СП № 5047-89 «Санитарные правила по гигиене труда для обувных предприятий»
СП № 5181-90 «Санитарные правила для производства полупроводниковых приборов и интегральных микросхем»
СП № 5182-90 «Санитарные правила для швейного производства»
СП № 5206-90 «Санитарные правила для катализаторных производств нефтеперерабатывающей и нефтехимической промышленности»
СП № 5312-91 «Санитарные правила для предприятий медно-никелевой промышленности»
СП № 6035-91 «Санитарные правила по устройству и эксплуатации средств малой механизации для сельскохозяйственного производства»
СП № 4950-89 «Санитарные правила для производств материалов на основе углерода (угольных, графитированных, волокнистых, композиционных) «
СП № 2527-82 «Санитарные правила для предприятий черной металлургии»
СП № 1889-78 «Санитарные правила по устройству и содержанию предприятий кожевенной промышленности»
СП № 1122-73 «Санитарные правила организации работы по напылению жесткого пенополиуретана»
СП № 1055-73 «Временные санитарные правила по устройству, оборудованию и содержанию предприятий по производству синтомицина и левомицетина»
СП № 984-72 «Санитарные правила по устройству, оборудованию и содержанию предприятий, изготовляющих люминофоры и люминесцентные лампы»
СП № 338-60 «Санитарные правила по устройству, оборудованию и содержанию предприятий по производству кремнийорганических соединений (алкил-, арилхлорсиланов) «
СП 2. 2.9.2510-09 «Гигиенические требования к условиям труда инвалидов»
2.9.2510-09 «Гигиенические требования к условиям труда инвалидов»
СП 2.5.1.1107-02 «Гигиенические требования к условиям и организации труда диспетчеров по управлению воздушным движением гражданской авиации»
СП № 6030-91 «Санитарные правила для работы с источниками ионизирующего излучения при обслуживании и ремонте воздушных судов на предприятиях и заводах гражданской авиации»
СП 2.5.1198-03 «Санитарных правил по организации пассажирских перевозок на железнодорожном транспорте»
СП 52.13330.2011 «Естественное и искусственное освещение. Актуализированная редакция СНиП 23-05-95*»
СП 44.13330.2011 «Административные и бытовые здания. Актуализированная редакция СНиП 2.09.04-87″
СП 118.13330.2012* «Общественные здания и сооружения. Актуализированная редакция СНиП 31-06-2009″
СП 158.13330.2014 «Здания и помещения медицинских организаций. Правила проектирования»
СП 131. 13330.2012 «Строительная климатология. Актуализированная редакция СНиП 23-01-99* «
13330.2012 «Строительная климатология. Актуализированная редакция СНиП 23-01-99* «
СП 60.13330.2012 «Отопление, вентиляция и кондиционирование воздуха. Актуализированная редакция СНиП 41-01-2003″
СП 2.5.1337-03 «Санитарные правила эксплуатации метрополитенов»
СП 2.1.2.3304-15 «Санитарно-эпидемиологические требования к размещению, устройству и содержанию объектов спорта»
СП 2.1.2.3358-16 «Санитарно- эпидемиологические требования к размещению, устройству, оборудованию, содержанию, санитарно-гигиеническому и противоэпидемическому режиму работы организаций социального обслуживания»
СП 2.6.1.3247-15 «Гигиенические требования к размещению, устройству, оборудованию и эксплуатации радоновых лабораторий, отделений радонотерапии»
СП 2.2.1.2263-07 «Санитарные правила для автотранспортного предприятия с топливозаправочным пунктом, осуществляющего заправку и эксплуатацию автомобилей на диметиловом эфире»
СП 2. 3.3.2892-11 «Санитарно-гигиенические требования к организации и проведению работ с метанолом»
3.3.2892-11 «Санитарно-гигиенические требования к организации и проведению работ с метанолом»
СП 5199-90 «Санитарные правила при производстве синтетических моющих средств»
СП 5183-90 «Санитарные правила для литейного производства (заводов, цехов, участков) «
СП 946-А-71 «Санитарные правила для предприятий маргариновой промышленности»
СП 962-72 «Санитарные правила для предприятий, вырабатывающих плодоовощные консервы, сушеные фрукты, овощи и картофель, квашеную капусту и соленые овощи»
СП 977-72 «Санитарные правила для предприятий чайной промышленности»
СП 989-72 «Санитарные правила для предприятий макаронной промышленности»
СП 1197-74 «Санитарные правила для предприятий по производству растительных масел
Санитарные правила для предприятий желатиновой промышленности (СП от 26.10.1978) «
СП 2266-80 «Санитарные правила для предприятий дрожжевой промышленности»
СП 2982-84 «Санитарные правила для предприятий по производству быстрозамороженных готовых блюд»
СП 3238-85 «Санитарные правила для предприятий мясной промышленности»
СП 3244-85 «Санитарные правила для предприятий пивоваренной и безалкогольной промышленности»
СП 4261-87 «Ветеринарно-санитарные правила для предприятий (цехов) переработки птицы и производства яйцепродуктов»
СП 5788-91″Санитарные правила для винодельческих предприятий»
СП 2. 3.4.009-93 «Санитарные правила по заготовке, переработке и продаже грибов»
3.4.009-93 «Санитарные правила по заготовке, переработке и продаже грибов»
СП 2.1.2.2844-11 «Санитарно-эпидемиологические требования к устройству, оборудованию и содержанию общежитий для работников организаций и обучающихся образовательных учреждений»
СП 3.3.2342-08 «Обеспечение безопасности иммунизации»
СП 3.3.2.3332-16 «Условия транспортирования и хранения иммунобиологических лекарственных препаратов»
СП 2.3.6.1079-01 «Санитарно-эпидемиологические требования к организациям общественного питания, изготовлению и оборотоспособности в них пищевых продуктов и продовольственного сырья»
СП 2.3.6.1066-01″Санитарно-эпидемиологические требования к организациям торговли и обороту в них продовольственного сырья и пищевых продуктов»
СП 2.6.1.2612-10 «Основные санитарные правила обеспечения радиационной безопасности (ОСПОРБ 99/2010)»
СП 2.6.1.2216-07 «Санитарно-защитные зоны и зоны наблюдения радиационных объектов. Условия эксплуатации и обоснование границ»
Условия эксплуатации и обоснование границ»
СП 1.3.2322-08 «Безопасность работы с микроорганизмами III-IV групп патогенности (опасности) и возбудителями паразитарных болезней»
СП 2.1.7.1038-01 «Гигиенические требования к устройству и содержанию полигонов для твердых бытовых отходов»
СП 5190-90 «Временные санитарные правила для производств литиевых химических источников тока различных электрохимических систем»
Указания к проектированию и эксплуатации установок искусственного ультрафиолетового облучения на промышленных предприятиях (24 мая 1974)
СП 2.6.1.2622-10 «Гигиенические требования по обеспечению радиационной безопасности на объектах хранения газового конденсата в подземных резервуарах, образованных с применением ядерно-взрыной технологии»
СП 2.6.1.3241-14 «Гигиенические требования по обеспечению радиационной безопасности при радионуклидной дефектоскопии»
СП 3. 5.1378-03 «Санитарно-эпидемиологические требования к организации и осуществлению дезинфекционной деятельности»
5.1378-03 «Санитарно-эпидемиологические требования к организации и осуществлению дезинфекционной деятельности»
СП 2.1.7.1386-03 «Санитарные правила по определению класса опасности токсичных отходов производства и потребления»
СП № 4060-85 «Лечебные пляжи. Санитарные правила устройства, оборудования и эксплуатации»
СП № 983-72 «Санитарные правила устройства и содержания общественных уборных»
СП 53.13330.2011 «Планировка и застройка территорий садоводческих (дачных) объединений граждан, здания и сооружения » Актуализированная редакция СНиП 30-02-97*
СП 118.13330.2012* «Общественные здания и сооружения. Актуализированная редакция СНиП 31-06-2009″
СП 3.5.3.3223-14 «Санитарно-эпидемиологические требования к организации и проведению дератизационных мероприятий»
СП 54.13330.2011 «Здания жилые многоквартирные. Актуализированная редакция СНиП 31-01-2003″
СП 3. 1.1.2521-09 «Профилактика холеры. Общие требования к эпидемиологическому надзору за холерой на территории Российской Федерации»
1.1.2521-09 «Профилактика холеры. Общие требования к эпидемиологическому надзору за холерой на территории Российской Федерации»
СП 2.5.3157-14 «Санитарно-эпидемиологические требования к перевозке железнодорожным транспортом организованных групп детей»
СП 3.1.3263-15 «Профилактика инфекционных заболеваний при эндоскопических вмешательствах»
СП 3.1.3310-15 «Профилактика инфекций, передающихся иксодовыми клещами»
СП 3.1.1.3473-17 «Профилактика брюшного тифа и паратифов»
СП № 2449-81 «Санитарные правила для предприятий соляной промышленности»
СП № 1408-76 «Санитарные правила для предприятий пищеконцентратной промышленности»
СП № 4416-87 «Санитарные правила для предприятий по обработке и розливу питьевых минеральных вод»
СП 1145-74 «Санитарные правила для предприятий по производству пищевых кислот»
СП № 942-71 «Санитарные правила для детских молочных кухонь»
СП 2. 1.5.1059-01 «Гигиенические требования к охране подземных вод от загрязнения»
1.5.1059-01 «Гигиенические требования к охране подземных вод от загрязнения»
СП 2.1.4.2625-10 «Зоны санитарной охраны источников питьевого водоснабжения г. Москвы»
СП 2.1.7.2570-10 Изменение N 1 в СП 2.1.7.1386-03 «Санитарные правила по определению класса опасности токсичных отходов производства и потребления»
СП 2.1.7.2850-11 «Изменения и дополнения N 2 к СП 2.1.7.1386-03 «Санитарные правила по определению класса опасности токсичных отходов производства и потребления»
СН 4249-87 «Санитарные нормы вибрации в кабине машиниста тягового подвижного состава железнодорожного транспорта»
СН 2.2.4/2.1.8.562-96 «Шум на рабочих местах, в помещениях жилых, общественных зданий и на территории жилой застройки. Санитарные нормы»
СН 2.2.4/2.1.8.566-96 «Производственная вибрация, вибрация в помещениях жилых и общественных зданий. Санитарные нормы»
СН 2.2.4/2. 1.8.583-96 «Инфразвук на рабочих местах, в жилых и общественных помещениях и на территории жилой застройки»
1.8.583-96 «Инфразвук на рабочих местах, в жилых и общественных помещениях и на территории жилой застройки»
СН N 3057-84 «Санитарные нормы допустимого шума, создаваемого изделиями медицинской техники в помещениях лечебно-профилактических учреждений»
ГН 2.1.6.3537-18 «Предельно допустимые концентрации (ПДК) микроорганизмов-продуцентов, бактериальных препаратов и их компонентов в атмосферном воздухе городских и сельских поселений» и гигиенических нормативов ГН 2.2.6.3538-18 «Предельно допустимые концентрации (ПДК) микроорганизмов-продуцентов, бактериальных препаратов и их компонентов в воздухе рабочей зоны»
ГН 2.1.6.3492-17 «Предельно допустимые концентрации (ПДК) загрязняющих веществ в атмосферном воздухе городских и сельских поселений» (с изменениями на 31 мая 2018 года)
ГН 2.1.2/2.2.1.1009-00 «Перечень асбестоцементных материалов и конструкций, разрешенных к применению в строительстве»
ГН. 2.1.8/2.2.4.2262-07 «Предельно допустимые уровни магнитных полей частотой 50 Гц в помещениях жилых, общественных зданий и на селитебных территориях»
2.1.8/2.2.4.2262-07 «Предельно допустимые уровни магнитных полей частотой 50 Гц в помещениях жилых, общественных зданий и на селитебных территориях»
ГН 2.2.5.2557-09 «Предельно допустимые уровни (ПДУ) загрязнения мышьяком поверхностей технологического оборудования и строительных конструкций производственных помещений»
ГН 2.2.5.2308-07 «Ориентировочные безопасные уровни воздействия (ОБУВ) вредных веществ в воздухе рабочей зоны»
ГН 2.1.6.2309-07 «Ориентировочные безопасные уровни воздействия (ОБУВ) загрязняющих веществ в атмосферном воздухе населенных мест»
ГН 1.2.3539-18 «Гигиенические нормативы содержания пестицидов в объектах окружающей среды (перечень)»
ГН 2.2.5.3532-18 «Предельно допустимые концентрации (ПДК) вредных веществ в воздухе рабочей зоны»
ГН 2.6.1.2159-07 «Содержание техногенных радионуклидов в металлах»
ГН 2.1.5.1315-03 «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
ГН 2. 1.5.2280-07 «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
1.5.2280-07 «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
ГН 2.1.5.1315-03 «Предельно допустимые концентрации (ПДК) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
ГН 2.1.7.2511-09 «Ориентировочно допустимые концентрации (ОДК) химических веществ в почве»
ГН 2.2.5.2893-11 «Предельно допустимые уровни (ПДУ) загрязнения кожных покровов вредными веществами»
ГН 2.3.3.972-00 «Предельно допустимые количества химических веществ, выделяющихся из материалов, контактирующих с пищевыми продуктами»
ГН 2.1.7.2041-06 «Предельно допустимые концентрации (ПДК) химических веществ в почве»
ГН 2.1.5.2312-08 «Ориентировочные допустимые уровни (ОДУ) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования. Дополнение N 1 к ГН 2.1.5.2307-07″
Дополнение N 1 к ГН 2.1.5.2307-07″
ГН 2.1.5.2307-07 «Ориентировочные допустимые уровни (ОДУ) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
ГН 2.1.5.2415-08 «Ориентировочные допустимые уровни (ОДУ) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования. Дополнение N 2 к ГН 2.1.5.2307-07″
ГН 2.1.5.2702-10 «Дополнение N 3 к ГН 2.1.5.2307-07 «Ориентировочные допустимые уровни (ОДУ) химических веществ в воде водных объектов хозяйственно-питьевого и культурно-бытового водопользования»
ГН 1.1.701-98 «Гигиенические критерии для обоснования необходимости разработки ПДК и ОБУВ (ОДУ) вредных веществ в воздухе рабочей зоны, атмосферном воздухе населенных мест, воде водных объектов»
ГН 2.1.6.2563-09 «Ориентировочные безопасные уровни воздействия (ОБУВ) отравляющих веществ кожно-нарывного действия в атмосферном воздухе населенных мест»
ГН 2. 1.6.3306-15 «Ориентировочный безопасный уровень воздействия (ОБУВ) O-изобутил-бета-N-диэтиламиноэтилтиолового эфира метилфосфоновой кислоты (вещества типа Vx) в атмосферном воздухе населенных мест и в зонах защитных мероприятий объектов по хранению и уничтожению химического оружия»
1.6.3306-15 «Ориентировочный безопасный уровень воздействия (ОБУВ) O-изобутил-бета-N-диэтиламиноэтилтиолового эфира метилфосфоновой кислоты (вещества типа Vx) в атмосферном воздухе населенных мест и в зонах защитных мероприятий объектов по хранению и уничтожению химического оружия»
ГН 2.1.6.2736-10 «Ориентировочный безопасный уровень воздействия (ОБУВ) О-изопропилметилфторфосфоната (зарина) в атмосферном воздухе населенных мест»
ГН 2.1.6.2556-09 «Ориентировочный безопасный уровень воздействия (ОБУВ) 2-хлорвиниларсиноксида (оксида люизита) в атмосферном воздухе населенных мест»
СНиП 23-05-95* «Естественное и искусственное освещение»
СНиП 2.10.02-84 «Здания и помещения для хранения и переработки сельскохозяйственной продукции»
СНиП 41-01-2003 «Отопление, вентиляция и кондиционирование»
СНиП 23-01-99* «Строительная климатология»
СНиП 31-04-2001 «Складские здания»
СНиП 31-01-2003 «Здания жилые многоквартирные»
ГОСТ 12. 2.032-78 «Система стандартов безопасности труда (ССБТ). Рабочее место при выполнении работ сидя. Общие эргономические требования»
2.032-78 «Система стандартов безопасности труда (ССБТ). Рабочее место при выполнении работ сидя. Общие эргономические требования»
ГОСТ 12.2.049-80 «Система стандартов безопасности труда (ССБТ). Оборудование производственное. Общие эргономические требования»
ГОСТ 17228-2014 «Самолеты пассажирские и транспортные. Допустимые уровни шума, создаваемые на местности»
ГОСТ 30494-2011 «Здания жилые и общественные. Параметры микроклимата в помещениях»
МУ 2.6.1.1892-04 «Гигиенические требования по обеспечению радиационной безопасности при проведении радионуклидной диагностики с помощью радиофармпрепаратов»
МУ 2.6.1.2135-06 «Гигиенические требования по обеспечению радиационной безопасности при лучевой терапии закрытыми радионуклидными источниками»
МУ 2.6.1.2797-10 Изменение 1 к МУ 2.6.1.2135-06 «Гигиенические требования по обеспечению радиационной безопасности при лучевой терапии закрытыми радионуклидными источниками»
МУ 2. 6.1.2500-09 «Организация надзора за обеспечением радиационной безопасности и проведение радиационного контроля в подразделении радионуклидной диагностики»
6.1.2500-09 «Организация надзора за обеспечением радиационной безопасности и проведение радиационного контроля в подразделении радионуклидной диагностики»
МУ 2.6.1.2808-10 «Обеспечение радиационной безопасности при проведении радионуклидной диагностики методами радиоиммунного анализа «in vitro»
МУ 2.6.1.2712-10 «Гигиенические требования по обеспечению радиационной безопасности при внутритканевой лучевой терапии (брахитерапии) методом имплантации закрытых радионуклидных источников»
МУ 3.5.1.3439-17 «Оценка чувствительности к дезинфицирующим средствам икроорганизмов, циркулирующих в медицинских организациях»
МУ 3.1.3420-17 «Обеспечение эпидемиологической безопасности нестерильных эндоскопических вмешательств на желудочно-кишечном тракте и дыхательных путях»
МУ 3.1.3490-17 «Изучение популяционного иммунитета к гриппу у населения Российской Федерации»
МУ 3.1.3342-16 «Эпидемиологический надзор за ВИЧ-инфекцией»
МУ 2. 1.10.3014-12 «Оценка радиационного риска у населения за счет длительного равномерного техногенного облучения в малых дозах»
1.10.3014-12 «Оценка радиационного риска у населения за счет длительного равномерного техногенного облучения в малых дозах»
МУ 2.6.1.2398-08 «Радиационный контроль и санитарно-эпидемиологическая оценка земельных участков под строительство жилых домов, зданий и сооружений общественного и производственного назначения в части обеспечения радиационной безопасности»
МУ 2.6.1.2222-07 «Прогноз доз облучения населения радионуклидами цезия и стронция при их попадании в окружающую среду»
МУ 2.6.1.2153-06 «Оперативная оценка доз облучения населения при радиоактивном загрязнении территории воздушным путем»
МУ 2.1.4.1184-03 «Методические указания по внедрению и применению санитарно-эпидемиологических правил и нормативов СанПиН 2.1.4.1116-02 «Питьевая вода. Гигиенические требования к качеству воды, расфасованной в емкости. Контроль качества»
МУ 2.6.1.2719-10 «Радиационный контроль и гигиеническая оценка источников питьевого водоснабжения и питьевой воды по показателям радиационной безопасности. Оптимизация защитных мероприятий источников питьевого водоснабжения с повышенным содержанием радионуклидов» Изменение N 1 к МУ 2.6.1.1981-05
Оптимизация защитных мероприятий источников питьевого водоснабжения с повышенным содержанием радионуклидов» Изменение N 1 к МУ 2.6.1.1981-05
МУ 2.6.1.1981-05 «Радиационный контроль и гигиеническая оценка источников питьевого водоснабжения и питьевой воды по показателям радиационной безопасности. Оптимизация защитных мероприятий источников питьевого водоснабжения с повышенным содержанием радионуклидов»
МУ 2.2.9.2493-09 «Санитарно-гигиеническая паспортизация канцерогеноопасных организаций и формирование банков данных»
МР 3.5.1.0100-15 «Применение установок импульсного ультрафиолетового излучения сплошного спектра в медицинских организациях»
МР 2.2.4.0115-16 «Оценка безопасности использования лазерных проекторов»
МР 2.3.1.1915-04 «Методические рекомендации. Рекомендуемые уровни потребления пищевых и биологически активных веществ»
МР 2.6.1.0091-14 «Радиационный контроль и санитарно-эпидемиологическая оценка минеральных удобрений и агрохимикатов по показателям радиационной безопасности»
МР 3. 5.1.0103-15 «Методические рекомендации по использованию метода аэрозольной дезинфекции в медицинских организациях»
5.1.0103-15 «Методические рекомендации по использованию метода аэрозольной дезинфекции в медицинских организациях»
Р 3.5.1904-04 «Использование ультрафиолетового бактерицидного излучения для обеззараживания воздуха в помещениях»
Р 2.2.4/2.2.9.2266-07 «Гигиенические требования к условиям труда медицинских работников, выполняющих ультразвуковые исследования»Технология стальных труб. Требования к устройству и эксплуатации взрывоопасного и химически опасного производства Язык: английский | Металлоконструкции Язык: английский | Нагрузки и действия Язык: английский | Сантехника керамическая. Язык: английский | Сосуды и аппараты стальные сварные. Общие технические условия Язык: английский | Прокат из высокопрочной стали. Общие технические условия Язык: английский | Знак соответствия формы обязательной сертификации, габаритов и технических требований Язык: английский | Сосуды и аппараты стальные сварные.Основные Характеристики. Язык: английский | Трубопроводная арматура. Скорость утечки клапанов. Язык: английский | Сварные стыковые соединения трубопроводов АЭС. Язык: английский | Сварные швы для стыков трубопроводов ТЭЦ.Типы и основные размеры. Язык: английский | Детали и узлы трубопроводов атомных станций из сталей перлитного класса на давление до 2,2 МПа (22 кгс / см2). Сварные швы. Типы и размеры Язык: английский | Технический регламент Таможенного союза. Безопасность высокоскоростного железнодорожного транспорта Язык: английский | Технический регламент Таможенного союза.О безопасности упаковки Язык: английский | Дуговая сварка в защитном газе. Язык: английский | Сталь и сплавы. Методы обнаружения и определения крупности Язык: английский | Фильтры жидкости периодического действия, работающие под давлением.Требования безопасности и методы испытаний Язык: английский | Масла вакуумные. Метод определения давления пара и температуры кипения Язык: английский | Скалы. Методы определения прочности на осевое сжатие Язык: английский | Основные нормы взаимозаменяемости. Язык: английский |
 Типы и габаритные размеры
Типы и габаритные размеры Типы и основные размеры
Типы и основные размеры Сварные соединения. Основные типы, элементы конструкции и размеры
Сварные соединения. Основные типы, элементы конструкции и размеры Общие допуски. Допуски формы и положения для элементов без указания индивидуальных допусков
Общие допуски. Допуски формы и положения для элементов без указания индивидуальных допусковЭтот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.

- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или уточнить у системного администратора.

Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файлах cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Европейский журнал полимеров — Elsevier
European Polymer Journal предназначен для публикации работ по фундаментальной и прикладной химии полимеров и макромолекулярных материалов. Журнал охватывает все аспекты синтеза полимеров, включая механизмы полимеризации и химические функциональные превращения, с акцентом на новые полимеры и. ..
..
European Polymer Journal предназначен для публикации работ по фундаментальной и прикладной химии полимеров и макромолекулярных материалов. Журнал охватывает все аспекты синтеза полимеров, включая механизмы полимеризации и химические функциональные превращения, с акцентом на новые полимеры и взаимосвязь между молекулярной структурой и свойствами полимера. Кроме того, мы приветствуем заявки на биологические или возобновляемые полимеры, системы, реагирующие на раздражители, и полимерные биогибриды. European Polymer Journal также публикует исследования по биомедицинскому применению полимеров, включая доставку лекарств и регенеративную медицину. Основная область охвата, но не ограничивается следующими основными областями исследований:
Синтез и функционализация полимеров
• Новые пути синтеза для полимеризации, функциональной модификации, контролируемой / живой полимеризации и прецизионных полимеров.
Полимеры, реагирующие на раздражение
• Включая полимеры с памятью формы и самовосстанавливающиеся полимеры.
Супрамолекулярные полимеры и самосборка
• Молекулярное распознавание и полимерные структуры более высокого порядка.
Возобновляемые и устойчивые полимеры
• Биоразлагаемые, антимикробные полимеры и полимерные бионокомпозиты на биологической основе.
Полимеры на границах раздела и поверхностях
• Химия и инженерия поверхностей, имеющих биологическое значение, включая формирование рисунка, полимеры против обрастания и полимеры для мембран.
Биомедицинские приложения и наномедицина
• Полимеры для регенеративной медицины, молекулярного высвобождения лекарств и генной терапии
В европейский журнал полимеров больше не входит физика полимеров.Мы настоятельно рекомендуем авторам рассмотреть возможность отправки статей по физике полимеров в наше отличное родственное название Polymer https://www.journals.elsevier.com/polymer
Типы статей и подача
European Polymer Journal публикует следующие типы статей: оригинальные исследования документы, обзорные статьи, избранные статьи, короткие сообщения и исследования. Пожалуйста, обратитесь к нашему Руководству для авторов для получения дополнительной информации о подаче статей. Если вам потребуется дополнительная информация или помощь, посетите наш Центр поддержки
Пожалуйста, обратитесь к нашему Руководству для авторов для получения дополнительной информации о подаче статей. Если вам потребуется дополнительная информация или помощь, посетите наш Центр поддержки
Преимущества для авторов
Мы также предоставляем множество преимуществ для авторов, такие как бесплатные PDF-файлы, либеральная политика авторских прав, специальные скидки на публикации Elsevier и многое другое.Щелкните здесь, чтобы получить дополнительную информацию о наших услугах для авторов.
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
 Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файлах cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
JoF | Бесплатный полнотекстовый | Микотоксины-аналоги сфинганина (SAM): химическая структура, биоактивность и генетический контроль
1.
 Введение Микотоксины — это вторичные метаболиты, продуцируемые различными грибами. Эти метаболиты выполняют важные экологические функции в живых системах в их естественной среде обитания. В качестве вторичных метаболитов микотоксины считаются несущественными для роста или размножения грибов. Однако их токсическое воздействие на растения, животных, а также людей привлекает все большее внимание химиков, биологов, ученых-диетологов и медицинских работников.Многие грибы способны синтезировать микотоксины, в том числе некоторые сапрофитные плесени, ядовитые грибы, грибковые патогены человека и грибковые патогены растений. Микотоксины, продуцируемые патогенными грибами растений, можно разделить на две группы: (i) селективные (или специфичные для хозяина) токсины (HST) и (ii) неспецифические токсины для хозяина (nHST), в зависимости от того, являются ли они специфически токсичными. для растения-хозяина (HST) или для широкого круга видов (nHST). Известные микотоксины обычно представляют собой химические вещества с низкой молекулярной массой, но с различными структурами и способами действия.
Введение Микотоксины — это вторичные метаболиты, продуцируемые различными грибами. Эти метаболиты выполняют важные экологические функции в живых системах в их естественной среде обитания. В качестве вторичных метаболитов микотоксины считаются несущественными для роста или размножения грибов. Однако их токсическое воздействие на растения, животных, а также людей привлекает все большее внимание химиков, биологов, ученых-диетологов и медицинских работников.Многие грибы способны синтезировать микотоксины, в том числе некоторые сапрофитные плесени, ядовитые грибы, грибковые патогены человека и грибковые патогены растений. Микотоксины, продуцируемые патогенными грибами растений, можно разделить на две группы: (i) селективные (или специфичные для хозяина) токсины (HST) и (ii) неспецифические токсины для хозяина (nHST), в зависимости от того, являются ли они специфически токсичными. для растения-хозяина (HST) или для широкого круга видов (nHST). Известные микотоксины обычно представляют собой химические вещества с низкой молекулярной массой, но с различными структурами и способами действия. Одна группа микотоксинов структурно аналогична сфингозину, основному предшественнику сфинголипидов, которые играют важные структурные и клеточные роли в эукариотических клетках. Эти токсины называются микотоксинами-аналогами сфинганина (SAM), вместе с фумонизинами и Alternaria alternata f. sp. Токсины Lycopersici (AAL) как две наиболее широко изученные группы SAM. SAM токсичны для растений и животных. Они действуют путем ингибирования церамидсинтазы (CerS), тем самым влияя на метаболизм сфинголипидов и инициируя апоптоз у животных и программируемую гибель клеток (PCD) у растений [1,2,3].Цель данной статьи — представить обновленный обзор структурного разнообразия, синтезов, способов действия и воздействия на здоровье SAM. Об открытии фумонизина впервые было сообщено в 1988 году, и организмом, продуцирующим его, был Fusarium verticillioides (син. Gibberella fujikuroi спаривающаяся популяция A, син. G. moniliformis Wineland, син. F. moniliforme Sheldon) [4]. С тех пор было обнаружено, что фумонизины продуцируются по крайней мере 18 видами рода Fusarium, наиболее заметными из которых являются F.
Одна группа микотоксинов структурно аналогична сфингозину, основному предшественнику сфинголипидов, которые играют важные структурные и клеточные роли в эукариотических клетках. Эти токсины называются микотоксинами-аналогами сфинганина (SAM), вместе с фумонизинами и Alternaria alternata f. sp. Токсины Lycopersici (AAL) как две наиболее широко изученные группы SAM. SAM токсичны для растений и животных. Они действуют путем ингибирования церамидсинтазы (CerS), тем самым влияя на метаболизм сфинголипидов и инициируя апоптоз у животных и программируемую гибель клеток (PCD) у растений [1,2,3].Цель данной статьи — представить обновленный обзор структурного разнообразия, синтезов, способов действия и воздействия на здоровье SAM. Об открытии фумонизина впервые было сообщено в 1988 году, и организмом, продуцирующим его, был Fusarium verticillioides (син. Gibberella fujikuroi спаривающаяся популяция A, син. G. moniliformis Wineland, син. F. moniliforme Sheldon) [4]. С тех пор было обнаружено, что фумонизины продуцируются по крайней мере 18 видами рода Fusarium, наиболее заметными из которых являются F. verticillioides и F. proliferatum, а также тремя неродственными родами грибов, секцией Aspergillus Nigri (такими как Asp.niger, Asp. Welwitschiae (син. Asp. Awamori) и т. Д., Известные как черные аспергиллы), Tolypocladium (T. inflatum, T. cylindrosporum и T.gedes) и Alternaria (патотип A. alternata, ранее известный как A. arborescens) [5,6,7,8,9,10]. Виды рода Fusarium можно найти как сапрофиты в почве, а также как эндофиты и патогены многих растений по всему миру. Распространенной группой болезней, вызываемых патогенами Fusarium, является гниение, которое может поражать все ткани на всех этапах развития растений [11,12].Кроме того, виды Fusarium могут инфицировать посевы в послеуборочный период при хранении [13]. Выжившие в почве пропагулы грибов также могут инфицировать новые культурные растения и могут быть перенесены на новые поля ветром или антропогенной деятельностью, например, при пересадке рассады [14]. Штаммы Fusarium могут синтезировать фумонизины на всех стадиях своего роста, включая сапрофитную стадию в почве, во время их патогенеза и в качестве эндофитов в различных частях растений, а также при хранении урожая после сбора урожая [15].
verticillioides и F. proliferatum, а также тремя неродственными родами грибов, секцией Aspergillus Nigri (такими как Asp.niger, Asp. Welwitschiae (син. Asp. Awamori) и т. Д., Известные как черные аспергиллы), Tolypocladium (T. inflatum, T. cylindrosporum и T.gedes) и Alternaria (патотип A. alternata, ранее известный как A. arborescens) [5,6,7,8,9,10]. Виды рода Fusarium можно найти как сапрофиты в почве, а также как эндофиты и патогены многих растений по всему миру. Распространенной группой болезней, вызываемых патогенами Fusarium, является гниение, которое может поражать все ткани на всех этапах развития растений [11,12].Кроме того, виды Fusarium могут инфицировать посевы в послеуборочный период при хранении [13]. Выжившие в почве пропагулы грибов также могут инфицировать новые культурные растения и могут быть перенесены на новые поля ветром или антропогенной деятельностью, например, при пересадке рассады [14]. Штаммы Fusarium могут синтезировать фумонизины на всех стадиях своего роста, включая сапрофитную стадию в почве, во время их патогенеза и в качестве эндофитов в различных частях растений, а также при хранении урожая после сбора урожая [15]. Фумонизины, как nHST, являются основными загрязнителями злаков и зерновых, включая кукурузу, рис, пшеницу, ячмень, рожь, овес, просо и продукты, изготовленные на основе этих культур [16]. Употребление продуктов питания, загрязненных фумонизинами, значительно увеличивает проблемы со здоровьем людей, что приводит к различным видам рака, таким как рак пищевода и неврологические дефекты [17,18]. Например, Международное агентство по изучению рака (IARC) охарактеризовало фумонизин FB 1 как канцерогены группы 2B для человека [16].Фумонизины также могут вызывать болезни и побочные эффекты у других видов, особенно у домашнего скота, когда корма загрязнены [19]. Хорошо известные болезни домашнего скота, вызываемые фумонизинами, включают лейкоэнцефаломаляцию у лошадей и синдром отека легких у свиней [20,21]. Подобно фуминизинам, AAL-токсины включают семейство структурно аналогичных метаболитов. AAL-токсины представляют собой группу HST, продуцируемых грибковым патогеном аскомицетов A.
Фумонизины, как nHST, являются основными загрязнителями злаков и зерновых, включая кукурузу, рис, пшеницу, ячмень, рожь, овес, просо и продукты, изготовленные на основе этих культур [16]. Употребление продуктов питания, загрязненных фумонизинами, значительно увеличивает проблемы со здоровьем людей, что приводит к различным видам рака, таким как рак пищевода и неврологические дефекты [17,18]. Например, Международное агентство по изучению рака (IARC) охарактеризовало фумонизин FB 1 как канцерогены группы 2B для человека [16].Фумонизины также могут вызывать болезни и побочные эффекты у других видов, особенно у домашнего скота, когда корма загрязнены [19]. Хорошо известные болезни домашнего скота, вызываемые фумонизинами, включают лейкоэнцефаломаляцию у лошадей и синдром отека легких у свиней [20,21]. Подобно фуминизинам, AAL-токсины включают семейство структурно аналогичных метаболитов. AAL-токсины представляют собой группу HST, продуцируемых грибковым патогеном аскомицетов A. alternata f. sp. Lycopersici, возбудитель язвы стебля томата [22].Следует отметить, что несколько других патотипов A. alternata могут также продуцировать другие HST, ответственные за грибковый патогенез на их конкретных растениях-хозяевах, соответственно [23]. В отличие от других HST, продуцируемых A. alternata, помимо восприимчивого хозяина томата, AAL-токсины также могут поражать многие другие сорняки и культуры двудольных видов и по крайней мере 25 видов Solanaceae [24,25]. Кроме того, патотип томатов A. alternata также вырабатывает фумонизин B (FB) [8,26]. AAL-токсин и FB были обнаружены не только в тканях некроза растений и культуральных средах, инокулированных A.alternata, но также в спорах и мицелиях этого патогена [27]. Однако AAL-токсин остается единственным токсином в качестве фактора патогенности для язвенной болезни стебля чувствительных сортов томатов, в то время как фумонизины являются токсигенными факторами вирулентности [28].
alternata f. sp. Lycopersici, возбудитель язвы стебля томата [22].Следует отметить, что несколько других патотипов A. alternata могут также продуцировать другие HST, ответственные за грибковый патогенез на их конкретных растениях-хозяевах, соответственно [23]. В отличие от других HST, продуцируемых A. alternata, помимо восприимчивого хозяина томата, AAL-токсины также могут поражать многие другие сорняки и культуры двудольных видов и по крайней мере 25 видов Solanaceae [24,25]. Кроме того, патотип томатов A. alternata также вырабатывает фумонизин B (FB) [8,26]. AAL-токсин и FB были обнаружены не только в тканях некроза растений и культуральных средах, инокулированных A.alternata, но также в спорах и мицелиях этого патогена [27]. Однако AAL-токсин остается единственным токсином в качестве фактора патогенности для язвенной болезни стебля чувствительных сортов томатов, в то время как фумонизины являются токсигенными факторами вирулентности [28]. Из-за неблагоприятного воздействия SAM на здоровье животных и человека эти токсины также привлекают все большее внимание инспекторов по контролю качества пищевых продуктов и органов общественного здравоохранения. За последние три десятилетия был достигнут значительный прогресс в нашем понимании SAM.Наши цели этого обзора — зафиксировать эти разработки SAM в отношении их химического структурного разнообразия, взаимосвязи между структурой и активностью, индукции PCD, детоксикации, генетики и эволюции биосинтеза SAM и лабораторных обнаружений.
За последние три десятилетия был достигнут значительный прогресс в нашем понимании SAM.Наши цели этого обзора — зафиксировать эти разработки SAM в отношении их химического структурного разнообразия, взаимосвязи между структурой и активностью, индукции PCD, детоксикации, генетики и эволюции биосинтеза SAM и лабораторных обнаружений.
3. Взаимосвязь между структурой SAM и биологической активностью
Биологические эффекты SAM, такие как их токсичность, одинаковы для разных SMA. Многие SAM имеют сходный спектр восприимчивых видов растений [34].Ткани и клетки томата одинаково чувствительны к AAL-токсинам и к токсинам FB 1 и FB 2 . У некоторых других растений AAL-токсины могут вызывать некротическую гибель клеток, аналогичную гибели фумонизинов [66]. Для культур тканей животных токсины ТА могут вызывать цитотоксичность как в печени крысы, так и в клетках почек собаки, как токсин FB 1 [67,68]. Кроме того, AAL-токсин и F. verticillioides могут также подавлять рост личинок и снижать вес куколок табачного почкоуборочного Heliothis virescens [69]. Такое сходство было приписано структурному сходству между SAM и сфинганином. Однако между SAM есть различия в их биологических эффектах, и эти различия связаны с их структурными различиями. Ниже мы суммируем основные результаты в этой области. Аминофункциональная группа SAM играет важную роль в их токсической активности. Перацетилированные производные AAL-токсинов и FB 1 являются биологически неактивными или имеют значительно сниженную токсичность как в биотестах растений, так и в системах культивирования тканей животных [66,70,71].Эти результаты согласуются с первоначальными сообщениями об этих токсинах, показывающими, что блокирование свободных первичных аминов AAL-токсинов специфическими реагентами может устранить биологическую активность этих токсинов в растениях [72]. В тесте первичных гепатоцитов крысы in vitro было отмечено, что N-ацетиловый аналог FB 1 , FA 1 также показал ингибирование CerS [68]. Позже было обнаружено, что FA спонтанно подвергается изомеризации, перестраивая свою O-ацетилированную группу с образованием других аналогов.
Такое сходство было приписано структурному сходству между SAM и сфинганином. Однако между SAM есть различия в их биологических эффектах, и эти различия связаны с их структурными различиями. Ниже мы суммируем основные результаты в этой области. Аминофункциональная группа SAM играет важную роль в их токсической активности. Перацетилированные производные AAL-токсинов и FB 1 являются биологически неактивными или имеют значительно сниженную токсичность как в биотестах растений, так и в системах культивирования тканей животных [66,70,71].Эти результаты согласуются с первоначальными сообщениями об этих токсинах, показывающими, что блокирование свободных первичных аминов AAL-токсинов специфическими реагентами может устранить биологическую активность этих токсинов в растениях [72]. В тесте первичных гепатоцитов крысы in vitro было отмечено, что N-ацетиловый аналог FB 1 , FA 1 также показал ингибирование CerS [68]. Позже было обнаружено, что FA спонтанно подвергается изомеризации, перестраивая свою O-ацетилированную группу с образованием других аналогов. Влияние этих продуктов реаранжировки на ингибирование CerS в срезах печени крыс также подтверждает важную роль первичной аминокислоты как в ингибировании, так и в токсичности CerS [73]. Дериватизация аминогруппы флуорогенными реагентами также делает возможным обнаружение FB с помощью анализа высокоэффективной жидкостной хроматографии (ВЭЖХ) [74]. FB могут ковалентно связываться с белками, вступая в реакцию с аминогруппами при абиогенных превращениях, что может повышать токсичность этих продуктов превращения [75].Точно так же терминальная аминогруппа FB 1 может конъюгироваться с бычьим сывороточным альбумином (BSA) и работать как иммуноген для получения моноклональных антител для обнаружения с помощью иммуноферментного анализа (ELISA) [76]. Аминогруппа фумонизинов также может работать как донор электронов и реагировать с электрофильным углеродом в изотиоцианатной (ITC) группе. Следовательно, FB могут быть разложены фумигационной обработкой ITC-содержащими соединениями [77]. Продукт гидролиза FB 1 (HFB 1 ) оказался менее токсичным, чем FB 1 и TA для растений [78] .
Влияние этих продуктов реаранжировки на ингибирование CerS в срезах печени крыс также подтверждает важную роль первичной аминокислоты как в ингибировании, так и в токсичности CerS [73]. Дериватизация аминогруппы флуорогенными реагентами также делает возможным обнаружение FB с помощью анализа высокоэффективной жидкостной хроматографии (ВЭЖХ) [74]. FB могут ковалентно связываться с белками, вступая в реакцию с аминогруппами при абиогенных превращениях, что может повышать токсичность этих продуктов превращения [75].Точно так же терминальная аминогруппа FB 1 может конъюгироваться с бычьим сывороточным альбумином (BSA) и работать как иммуноген для получения моноклональных антител для обнаружения с помощью иммуноферментного анализа (ELISA) [76]. Аминогруппа фумонизинов также может работать как донор электронов и реагировать с электрофильным углеродом в изотиоцианатной (ITC) группе. Следовательно, FB могут быть разложены фумигационной обработкой ITC-содержащими соединениями [77]. Продукт гидролиза FB 1 (HFB 1 ) оказался менее токсичным, чем FB 1 и TA для растений [78] . Ни HFB 1 , ни сфинголипиды дрожжей (полностью ацетилированные) не содержат PTCA. Хотя оба препарата оказали неблагоприятное воздействие на рост ряски, они показали более низкую фитотоксичность, чем TA и FB 1 , которые содержали один и два PTCA, соответственно [79]. Напротив, продукты гидролиза AAL-токсинов в значительной степени поддерживают токсичность своих исходных соединений для чувствительных линий томатов [66]. Эти результаты показывают, что PTCA важна для фитотоксичности FB и что существует специфичность взаимодействия между AAL-токсинами и томатами.В отличие от растений, тест in vitro с использованием первичных гепатоцитов крысы показал, что HFB обладают большей цитотоксичностью, чем FB. Однако HFB не могут инициировать развитие рака из-за отсутствия фрагмента PTCA, который, как предполагалось, играет активную роль в абсорбции фумонизинов из кишечника [70]. В модели беременных мышей LM / Bc HFB 1 не вызывал дефектов нервной трубки. Напротив, 10 мг FB 1 / кг массы тела мышей нарушили метаболизм материнских сфинголипидов, вызвали апоптоз печени у самок мышей, увеличили смертность плода и уменьшили вес плода [80].
Ни HFB 1 , ни сфинголипиды дрожжей (полностью ацетилированные) не содержат PTCA. Хотя оба препарата оказали неблагоприятное воздействие на рост ряски, они показали более низкую фитотоксичность, чем TA и FB 1 , которые содержали один и два PTCA, соответственно [79]. Напротив, продукты гидролиза AAL-токсинов в значительной степени поддерживают токсичность своих исходных соединений для чувствительных линий томатов [66]. Эти результаты показывают, что PTCA важна для фитотоксичности FB и что существует специфичность взаимодействия между AAL-токсинами и томатами.В отличие от растений, тест in vitro с использованием первичных гепатоцитов крысы показал, что HFB обладают большей цитотоксичностью, чем FB. Однако HFB не могут инициировать развитие рака из-за отсутствия фрагмента PTCA, который, как предполагалось, играет активную роль в абсорбции фумонизинов из кишечника [70]. В модели беременных мышей LM / Bc HFB 1 не вызывал дефектов нервной трубки. Напротив, 10 мг FB 1 / кг массы тела мышей нарушили метаболизм материнских сфинголипидов, вызвали апоптоз печени у самок мышей, увеличили смертность плода и уменьшили вес плода [80]. На модели свиней, чувствительных к SAMs, было показано, что HFB 1 имеет ограниченную кишечную или печеночную токсичность, но лишь незначительно нарушает метаболизм сфинголипидов [81]. Также были исследованы токсические эффекты воздействия FB 1 и HFB 1 на функцию кишечного барьера и иммунитет в модели совместной культуры кишечных эпителиальных клеток свиней и мононуклеарных клеток периферической крови свиней. FB 1 усиливает воспаление кишечника, вызванное липополисахаридом (LPS) / дезоксиниваленолом (DON), тогда как HFB 1 проявляет меньшую токсичность для иммунной системы [82].Кроме того, когда HFB 1 и HFB 2 были ацилированы с помощью CerS, N-ацил-метаболиты были токсичными in vitro для линии клеток толстой кишки человека и in vivo для внутрибрюшинных тканей крысы [83]. связывание с полисахаридами и белками через их две боковые цепи PTCA в термически обработанной пище и образование фумонизиновых артефактов [84].
На модели свиней, чувствительных к SAMs, было показано, что HFB 1 имеет ограниченную кишечную или печеночную токсичность, но лишь незначительно нарушает метаболизм сфинголипидов [81]. Также были исследованы токсические эффекты воздействия FB 1 и HFB 1 на функцию кишечного барьера и иммунитет в модели совместной культуры кишечных эпителиальных клеток свиней и мононуклеарных клеток периферической крови свиней. FB 1 усиливает воспаление кишечника, вызванное липополисахаридом (LPS) / дезоксиниваленолом (DON), тогда как HFB 1 проявляет меньшую токсичность для иммунной системы [82].Кроме того, когда HFB 1 и HFB 2 были ацилированы с помощью CerS, N-ацил-метаболиты были токсичными in vitro для линии клеток толстой кишки человека и in vivo для внутрибрюшинных тканей крысы [83]. связывание с полисахаридами и белками через их две боковые цепи PTCA в термически обработанной пище и образование фумонизиновых артефактов [84]. Активность SAM варьируется в зависимости от того, где происходит гидроксилирование углеродной основы. Например, FB 2 обладал более сильным цитотоксическим действием, чем FB 3 и FB 1 в первичных гепатоцитах крысы [70].Однако, в отличие от большинства других побочных групп, концевая метильная группа C-1, которая отличалась между FC и AAL-токсином от других фумонизинов, по-видимому, не требовалась для биологической активности SAM. Подобные симптомы, но меньшая фитотоксичность SAM наблюдались при длительном использовании -цепные сфингоидные основания или простые сфинголипиды были применены к ряске, что указывает на то, что фитотоксичность SAM может быть результатом накопления фитотоксических промежуточных сфинголипидов [71,85]. Этот результат соответствовал индукции PCD через сигнальные пути на основе церамидов (описанные ниже).Хотя AAL-токсины и фумонизины являются химическими веществами, родственными по структуре, с аналогичной фитотоксичностью, последние в 10 раз менее эффективны.
Активность SAM варьируется в зависимости от того, где происходит гидроксилирование углеродной основы. Например, FB 2 обладал более сильным цитотоксическим действием, чем FB 3 и FB 1 в первичных гепатоцитах крысы [70].Однако, в отличие от большинства других побочных групп, концевая метильная группа C-1, которая отличалась между FC и AAL-токсином от других фумонизинов, по-видимому, не требовалась для биологической активности SAM. Подобные симптомы, но меньшая фитотоксичность SAM наблюдались при длительном использовании -цепные сфингоидные основания или простые сфинголипиды были применены к ряске, что указывает на то, что фитотоксичность SAM может быть результатом накопления фитотоксических промежуточных сфинголипидов [71,85]. Этот результат соответствовал индукции PCD через сигнальные пути на основе церамидов (описанные ниже).Хотя AAL-токсины и фумонизины являются химическими веществами, родственными по структуре, с аналогичной фитотоксичностью, последние в 10 раз менее эффективны. Считается, что AAL-токсины служат гербицидом в очень низкой дозировке против широкого разнообразия широколистных сорняков (например, дурман, колючая сида и паслен черный). Однако однодольные культуры (например, кукуруза, пшеница и устойчивые сорта томатов) толерантны к AAL-токсинам [24,86,87]. До 2013 года механизм действия посредством ингибирования CerS не входил в число 21 молекулярного сайта-мишени обычно используемых гербицидов.Использование AAL-токсина в качестве ведущего соединения может создать новый и безопасный биогербицид, который обладает фитотоксичностью, но снижает или не токсичен для млекопитающих [88,89].
Считается, что AAL-токсины служат гербицидом в очень низкой дозировке против широкого разнообразия широколистных сорняков (например, дурман, колючая сида и паслен черный). Однако однодольные культуры (например, кукуруза, пшеница и устойчивые сорта томатов) толерантны к AAL-токсинам [24,86,87]. До 2013 года механизм действия посредством ингибирования CerS не входил в число 21 молекулярного сайта-мишени обычно используемых гербицидов.Использование AAL-токсина в качестве ведущего соединения может создать новый и безопасный биогербицид, который обладает фитотоксичностью, но снижает или не токсичен для млекопитающих [88,89].4. Детоксикация SAM
Использование сельскохозяйственных и производственных методов для предотвращения распространения и роста токсин-продуцирующих грибов и ограничения производства микотоксинов является предпочтительным методом устранения заражения пищевых продуктов грибковыми токсинами в предуборочный период [90]. Однако полностью предотвратить рост грибковых патогенов и заражение микотоксинами при ведении сельского хозяйства, а также при хранении и переработке пищевых продуктов крайне сложно. Поскольку токсичность SAM структурно зависит, наши знания о взаимосвязи между структурой SAM и биологической активностью дают подсказки для разработки эффективных стратегий управления для минимизации воздействия SAM на продукты питания и корма. Действительно, структурные модификации, такие как гидролиз, продемонстрировали свою эффективность в снижении токсичности SAM [91]. За последние три десятилетия были разработаны химические, биологические и физические стратегии для разложения микотоксинов в пищевых продуктах и кормах [92].Например, никстамализация применялась для уменьшения количества FB путем варки кукурузы, зараженной фумонизином, с известью, а также путем обработки атмосферным аммиаком [93,94,95]. Диоксид хлора также показал способность разлагать FB 1 [96]. Было показано, что два распространенных метода варки, включая экструзию и никстамализацию, снижают токсичность кукурузы, зараженной FB 1 [97]. Холодная плазма атмосферного давления использовалась в качестве физической обработки для успешного разложения чистых FB 1 и AAL-токсинов в течение 60 с, в то время как присутствие матрицы замедляло разложение [98,99].
Поскольку токсичность SAM структурно зависит, наши знания о взаимосвязи между структурой SAM и биологической активностью дают подсказки для разработки эффективных стратегий управления для минимизации воздействия SAM на продукты питания и корма. Действительно, структурные модификации, такие как гидролиз, продемонстрировали свою эффективность в снижении токсичности SAM [91]. За последние три десятилетия были разработаны химические, биологические и физические стратегии для разложения микотоксинов в пищевых продуктах и кормах [92].Например, никстамализация применялась для уменьшения количества FB путем варки кукурузы, зараженной фумонизином, с известью, а также путем обработки атмосферным аммиаком [93,94,95]. Диоксид хлора также показал способность разлагать FB 1 [96]. Было показано, что два распространенных метода варки, включая экструзию и никстамализацию, снижают токсичность кукурузы, зараженной FB 1 [97]. Холодная плазма атмосферного давления использовалась в качестве физической обработки для успешного разложения чистых FB 1 и AAL-токсинов в течение 60 с, в то время как присутствие матрицы замедляло разложение [98,99]. Озон применялся для разрушения клеток грибов Fusarium и Aspergillus путем окисления сульфгидрильных и аминокислотных групп ферментов или воздействия на полиненасыщенные жирные кислоты клеточной стенки [100]. Однако не все SAM подвержены физическим и химическим воздействиям. Кроме того, некоторые из этих обработок могут также приводить к получению производных с неизвестной токсичностью и быть вредными для обрабатываемых товаров, как показано в некоторых случаях [101, 102]. Другой метод снижения токсичности SAM — это воздействие микробов.Микроорганизмы могут проводить реакции биотрансформации для детоксикации SAM [91]. Такие методы включают дезаминирование, ацетилирование, гидролиз, глюкозилирование и декарбоксилирование. Например, Benedetti et al. изолировал штамм грамотрицательных палочек из почвы, способный расщеплять фумонизин до четырех метаболитов, когда фумонизин поставлялся в качестве единственного источника углерода [103]. Бактерия Sphingopyxis sp. может детоксифицировать фумонизин B1, по крайней мере, двумя ферментативными стадиями, включая начальную реакцию деэтерификации с последующим деаминированием гидролизованного продукта [104].
Озон применялся для разрушения клеток грибов Fusarium и Aspergillus путем окисления сульфгидрильных и аминокислотных групп ферментов или воздействия на полиненасыщенные жирные кислоты клеточной стенки [100]. Однако не все SAM подвержены физическим и химическим воздействиям. Кроме того, некоторые из этих обработок могут также приводить к получению производных с неизвестной токсичностью и быть вредными для обрабатываемых товаров, как показано в некоторых случаях [101, 102]. Другой метод снижения токсичности SAM — это воздействие микробов.Микроорганизмы могут проводить реакции биотрансформации для детоксикации SAM [91]. Такие методы включают дезаминирование, ацетилирование, гидролиз, глюкозилирование и декарбоксилирование. Например, Benedetti et al. изолировал штамм грамотрицательных палочек из почвы, способный расщеплять фумонизин до четырех метаболитов, когда фумонизин поставлялся в качестве единственного источника углерода [103]. Бактерия Sphingopyxis sp. может детоксифицировать фумонизин B1, по крайней мере, двумя ферментативными стадиями, включая начальную реакцию деэтерификации с последующим деаминированием гидролизованного продукта [104]. Chlebicz и likaewska обнаружили, что 12 штаммов Lactobacillus sp. бактерии и 6 штаммов дрожжей Saccharomyces cerevisiae могли снизить концентрацию FB 1 и FB 2 на 40% [105]. Аналогичным образом, Берджесс продемонстрировал, что продуцирующий фумонизин Asp. welwitschiae обладают способностью продуцировать ферменты для синтеза неаминированных фумонизинов, которые менее токсичны, чем FB, и эти ферменты могут использоваться для детоксикации фумонизина [106]. Действительно, использование ферментов для детоксикации путем модификации химической структуры стало многообещающим методом контроля микотоксинов после сбора урожая зерна [107,108].Например, fumD (карбоксилэстераза) от Sphingopyxis катализирует детоксикацию FB 1 до гидролизованной формы путем гидролиза обеих боковых цепей PTCA. Затем аминотрансфераза FumI могла разрушать FB 1 , катализируя дезаминирование HFB 1 . FumD также был протестирован на предмет влияния на адсорбцию фумонизинов у индейки, свиней и свиней [109,110,111].
Chlebicz и likaewska обнаружили, что 12 штаммов Lactobacillus sp. бактерии и 6 штаммов дрожжей Saccharomyces cerevisiae могли снизить концентрацию FB 1 и FB 2 на 40% [105]. Аналогичным образом, Берджесс продемонстрировал, что продуцирующий фумонизин Asp. welwitschiae обладают способностью продуцировать ферменты для синтеза неаминированных фумонизинов, которые менее токсичны, чем FB, и эти ферменты могут использоваться для детоксикации фумонизина [106]. Действительно, использование ферментов для детоксикации путем модификации химической структуры стало многообещающим методом контроля микотоксинов после сбора урожая зерна [107,108].Например, fumD (карбоксилэстераза) от Sphingopyxis катализирует детоксикацию FB 1 до гидролизованной формы путем гидролиза обеих боковых цепей PTCA. Затем аминотрансфераза FumI могла разрушать FB 1 , катализируя дезаминирование HFB 1 . FumD также был протестирован на предмет влияния на адсорбцию фумонизинов у индейки, свиней и свиней [109,110,111]. Наконец, некоторые другие ферменты, такие как пероксидаза марганца из разрушающих лигноцеллюлозу грибов и лакказа из Pleurotus eryngii, были способны разлагать фумонизины [112,113].Еще один потенциальный метод уменьшения количества SAM из пищевых продуктов и кормов — это использование адсорбирующих материалов для поглощения и удаления токсинов. Многие материалы продемонстрировали способность адсорбировать микотоксины in vitro, поэтому использование адсорбентов в рационе домашнего скота в качестве кормовых добавок может потенциально снизить биодоступность микотоксинов для человека и животных. В качестве кормовых добавок холестирамин, пластинки наносиликатной глины и очищенная кальциевая монтмориллонитовая глина снижали токсикоз FB 1 [114,115,116].Более того, натуральные продукты, такие как фенольное соединение хлорофорин, мед и коричное масло, оказались многообещающими в качестве агентов, снижающих уровень фумонизина [117,118,119].
Наконец, некоторые другие ферменты, такие как пероксидаза марганца из разрушающих лигноцеллюлозу грибов и лакказа из Pleurotus eryngii, были способны разлагать фумонизины [112,113].Еще один потенциальный метод уменьшения количества SAM из пищевых продуктов и кормов — это использование адсорбирующих материалов для поглощения и удаления токсинов. Многие материалы продемонстрировали способность адсорбировать микотоксины in vitro, поэтому использование адсорбентов в рационе домашнего скота в качестве кормовых добавок может потенциально снизить биодоступность микотоксинов для человека и животных. В качестве кормовых добавок холестирамин, пластинки наносиликатной глины и очищенная кальциевая монтмориллонитовая глина снижали токсикоз FB 1 [114,115,116].Более того, натуральные продукты, такие как фенольное соединение хлорофорин, мед и коричное масло, оказались многообещающими в качестве агентов, снижающих уровень фумонизина [117,118,119].5. Запрограммированная смерть клеток и сфинголипиды
Почти все клетки рано или поздно умирают. Существует четыре основных типа гибели клеток: некроптоз, пироптоз, ферроптоз и апоптоз, которые классифицируются на основе различных молекулярных и клеточных процессов и различных исходов. Апоптоз или запрограммированная смерть клеток (PCD) — это своего рода самоубийство клеток, которое строго регулирует клетки, которые больше не нужны или представляют угрозу для благополучия многоклеточных организмов.И у растений, и у животных есть PCD, и они функционально аналогичны друг другу [120,121,122,123]. PCD играет важную роль в поддержании нормальной физиологической активности многоклеточных организмов, таких как растения и животные, и представляет собой активный саморегулирующийся процесс для выборочного устранения избыточных, старых и поврежденных клеток. PCD можно предсказать для конкретных клеток на определенных стадиях развития. Однако PCD также может быть вызван мембраносвязанными и цитозольными белками, которые стимулируются сигналами, вызванными стрессом.Такие сигналы могут запускать гибель клеток посредством сложных каскадов транскрипционных изменений и посттрансляционных модификаций белков [122,124].
Существует четыре основных типа гибели клеток: некроптоз, пироптоз, ферроптоз и апоптоз, которые классифицируются на основе различных молекулярных и клеточных процессов и различных исходов. Апоптоз или запрограммированная смерть клеток (PCD) — это своего рода самоубийство клеток, которое строго регулирует клетки, которые больше не нужны или представляют угрозу для благополучия многоклеточных организмов.И у растений, и у животных есть PCD, и они функционально аналогичны друг другу [120,121,122,123]. PCD играет важную роль в поддержании нормальной физиологической активности многоклеточных организмов, таких как растения и животные, и представляет собой активный саморегулирующийся процесс для выборочного устранения избыточных, старых и поврежденных клеток. PCD можно предсказать для конкретных клеток на определенных стадиях развития. Однако PCD также может быть вызван мембраносвязанными и цитозольными белками, которые стимулируются сигналами, вызванными стрессом.Такие сигналы могут запускать гибель клеток посредством сложных каскадов транскрипционных изменений и посттрансляционных модификаций белков [122,124]. Характеристики PCD включают уменьшение объема клетки, маргинализацию и конденсацию хроматина, разборку ядерной ламины, фрагментацию ДНК, образование апоптотических тел и т. Д. [1]. Триггеры и распространение PCD включают множество факторов, включая экспрессию определенных рецепторов на поверхности клетки, трансмембранные домены нескольких мембранных белков, внутриклеточные белки, связанные с распространением сигналов смерти, вторичные мессенджеры, включая инозитолтрифосфат и церамиды, потоки кальция (Ca 2+ ), активные формы кислорода (ROS), регуляторные факторы клеточного цикла и другие супрессоры или активаторы белков.Многие из этих субклеточных компонентов, генов и путей передачи сигналов, участвующих в PCD, функционально законсервированы во всех доменах клеточных организмов, от бактерий до грибов, растений и животных. Однако существуют различия в фактических механизмах среди организмов, как обобщено в ссылках [125,126]. Предполагается, что сфинголипиды играют важную роль в росте, развитии клеток, реакции на внешнюю среду и PCD.
Характеристики PCD включают уменьшение объема клетки, маргинализацию и конденсацию хроматина, разборку ядерной ламины, фрагментацию ДНК, образование апоптотических тел и т. Д. [1]. Триггеры и распространение PCD включают множество факторов, включая экспрессию определенных рецепторов на поверхности клетки, трансмембранные домены нескольких мембранных белков, внутриклеточные белки, связанные с распространением сигналов смерти, вторичные мессенджеры, включая инозитолтрифосфат и церамиды, потоки кальция (Ca 2+ ), активные формы кислорода (ROS), регуляторные факторы клеточного цикла и другие супрессоры или активаторы белков.Многие из этих субклеточных компонентов, генов и путей передачи сигналов, участвующих в PCD, функционально законсервированы во всех доменах клеточных организмов, от бактерий до грибов, растений и животных. Однако существуют различия в фактических механизмах среди организмов, как обобщено в ссылках [125,126]. Предполагается, что сфинголипиды играют важную роль в росте, развитии клеток, реакции на внешнюю среду и PCD. Как главный компонент системы клеточных мембран, сфинголипиды помогают поддерживать структурную стабильность и транспорт молекул через клеточные мембраны [127,128].У млекопитающих сфинголипиды особенно распространены в клетках нервной системы, выполняя важные функции в контакте с клетками, росте, дифференцировке, коммуникации, ответе на сигналы стресса и апоптозе [30,129]. У растений сфинголипиды участвуют в реакции как на биотические, так и на абиотические стрессы, такие как инфекция патогенами, засуха и низкая температура [31,130,131]. Действительно, связь передачи сигналов церамидов с апоптозом широко известна как у растений, так и у животных. Следовательно, действия SAM по нарушению функций сфинголипидов могут иметь значительные негативные последствия.Однако наши знания о роли сфинголипидов в апоптозе также привели к растущему интересу к потенциальным новым методам лечения с использованием сфинголипидов в качестве мишеней лечения дегенеративных и пролиферативных заболеваний у людей и животных, таких как рак и болезнь Паркинсона [132, 133].
Как главный компонент системы клеточных мембран, сфинголипиды помогают поддерживать структурную стабильность и транспорт молекул через клеточные мембраны [127,128].У млекопитающих сфинголипиды особенно распространены в клетках нервной системы, выполняя важные функции в контакте с клетками, росте, дифференцировке, коммуникации, ответе на сигналы стресса и апоптозе [30,129]. У растений сфинголипиды участвуют в реакции как на биотические, так и на абиотические стрессы, такие как инфекция патогенами, засуха и низкая температура [31,130,131]. Действительно, связь передачи сигналов церамидов с апоптозом широко известна как у растений, так и у животных. Следовательно, действия SAM по нарушению функций сфинголипидов могут иметь значительные негативные последствия.Однако наши знания о роли сфинголипидов в апоптозе также привели к растущему интересу к потенциальным новым методам лечения с использованием сфинголипидов в качестве мишеней лечения дегенеративных и пролиферативных заболеваний у людей и животных, таких как рак и болезнь Паркинсона [132, 133]. исследования показали, что сфинголипиды могут служить важными вторичными посредниками в передаче сигналов для регуляции PCD [134,135]. Например, в нейтрофилах сфинголипиды связаны с повышенным образованием супероксида и притоком Ca 2+ , которые являются универсальными сигнальными молекулами, участвующими во многих клеточных функциях [136].Было также подтверждено, что индукция PCD у Arabidopsis церамидами частично зависит от ROS в митохондриях или регулируется высвобождением Ca 2+ [137,138]. Кроме того, повышение уровня сфингозина может активировать путь митоген-активируемой протеинкиназы (MAPK), в котором участвует MPK6, способствовать накоплению сиаловой кислоты (SA), а затем вызвать PCD [43]. Lachaud et al. обнаружили, что активированная кальций-зависимая киназа (CPK3) может регулировать процесс вызванной сфингозином PCD путем диссоциации CPK3 от белкового комплекса 14-3-3 при повышенной концентрации кальция, индуцированной сфингозином.Активированный CPK3 затем разлагается с последующей индукцией PCD [139].
исследования показали, что сфинголипиды могут служить важными вторичными посредниками в передаче сигналов для регуляции PCD [134,135]. Например, в нейтрофилах сфинголипиды связаны с повышенным образованием супероксида и притоком Ca 2+ , которые являются универсальными сигнальными молекулами, участвующими во многих клеточных функциях [136].Было также подтверждено, что индукция PCD у Arabidopsis церамидами частично зависит от ROS в митохондриях или регулируется высвобождением Ca 2+ [137,138]. Кроме того, повышение уровня сфингозина может активировать путь митоген-активируемой протеинкиназы (MAPK), в котором участвует MPK6, способствовать накоплению сиаловой кислоты (SA), а затем вызвать PCD [43]. Lachaud et al. обнаружили, что активированная кальций-зависимая киназа (CPK3) может регулировать процесс вызванной сфингозином PCD путем диссоциации CPK3 от белкового комплекса 14-3-3 при повышенной концентрации кальция, индуцированной сфингозином.Активированный CPK3 затем разлагается с последующей индукцией PCD [139]. Мутационные исследования Arabidopsis и эксперименты in vitro показали, что церамиды и свободные сфингоидные основания, такие как сфингозин, сфинганин и фитосфингозин, все могут вызывать PCD. Напротив, фосфорилированные продукты этих соединений могут ингибировать или ослаблять PCD у растений [44,138,140,141,142]. Однако фосфорилированный сфингозин может подавлять рост дрожжевых клеток, что позволяет предположить, что существуют разные механизмы действия между растениями и дрожжами [143].Стоит отметить, что не все керамиды могут вызывать PCD. Нагано и др. обнаружили, что когда положение C2 жирной кислоты в боковой цепи церамидов гидроксилируется, PCD скорее ингибируется, чем индуцируется [144]. Эти результаты показывают, что возникновение ПХД в растениях зависит не только от абсолютного содержания сфинголипидов, но и от относительных соотношений различных модифицированных форм. В следующем разделе мы опишем, как SAM участвуют в PCD.
Мутационные исследования Arabidopsis и эксперименты in vitro показали, что церамиды и свободные сфингоидные основания, такие как сфингозин, сфинганин и фитосфингозин, все могут вызывать PCD. Напротив, фосфорилированные продукты этих соединений могут ингибировать или ослаблять PCD у растений [44,138,140,141,142]. Однако фосфорилированный сфингозин может подавлять рост дрожжевых клеток, что позволяет предположить, что существуют разные механизмы действия между растениями и дрожжами [143].Стоит отметить, что не все керамиды могут вызывать PCD. Нагано и др. обнаружили, что когда положение C2 жирной кислоты в боковой цепи церамидов гидроксилируется, PCD скорее ингибируется, чем индуцируется [144]. Эти результаты показывают, что возникновение ПХД в растениях зависит не только от абсолютного содержания сфинголипидов, но и от относительных соотношений различных модифицированных форм. В следующем разделе мы опишем, как SAM участвуют в PCD.6. SAM запускают PCD через сигнальные пути на основе церамидов
В растениях вторжение патогенов может привести к нарушению гомеостаза клеток хозяина и вызвать гибель клеток у восприимчивых сортов или даже у устойчивых сортов с гиперчувствительным ответом (HR). Из-за большого сходства с PCD HR часто рассматривается как форма PCD у растений. AAL-токсины, являющиеся патогенным фактором язвенной болезни ствола томатов, могут вызывать PCD у чувствительных сортов томатов, что приводит к фрагментации хромосомной ДНК и образованию апоптотических телец в клетках [145]. Аналогичным образом, при обработке FB 1 , продуцируемым патогенным Fusarium, протопласты Arabidopsis проявляли симптомы, сходные с PCD в клетках животных [146]. На уровне тканей и органов листья арабидопсиса, обработанные токсином FB 1 , демонстрировали характерные симптомы заболевания.Клетки пораженных листьев имели общие фенотипы, сходные с HR, включая накопление каллозы, продукцию ROS и индукцию генов, связанных с патогенезом (PR) [147]. Повреждения, вызванные SAM на растениях-хозяевах, могут еще больше усилить инфекцию и колонизацию патогенов. Подобно тем, что обнаружены в растениях, SAM могут вызывать нейро- / почечные реакции, гептатоксикоз и новообразования, а также гибель клеток у животных.
Из-за большого сходства с PCD HR часто рассматривается как форма PCD у растений. AAL-токсины, являющиеся патогенным фактором язвенной болезни ствола томатов, могут вызывать PCD у чувствительных сортов томатов, что приводит к фрагментации хромосомной ДНК и образованию апоптотических телец в клетках [145]. Аналогичным образом, при обработке FB 1 , продуцируемым патогенным Fusarium, протопласты Arabidopsis проявляли симптомы, сходные с PCD в клетках животных [146]. На уровне тканей и органов листья арабидопсиса, обработанные токсином FB 1 , демонстрировали характерные симптомы заболевания.Клетки пораженных листьев имели общие фенотипы, сходные с HR, включая накопление каллозы, продукцию ROS и индукцию генов, связанных с патогенезом (PR) [147]. Повреждения, вызванные SAM на растениях-хозяевах, могут еще больше усилить инфекцию и колонизацию патогенов. Подобно тем, что обнаружены в растениях, SAM могут вызывать нейро- / почечные реакции, гептатоксикоз и новообразования, а также гибель клеток у животных. Взаимосвязь между апоптозом и передачей сигналов церамидов была установлена как у растений, так и у животных в их ответе на SAM [120].Например, индукция гибели клеток как в клетках почек томатов, так и в клетках почки африканской зеленой мартышки (CV-1) происходила при одинаковых концентрациях токсина и временных рамках. Для обоих типов клеток наблюдались морфологические маркеры, характерные для апоптоза, включая клетки с положительным концевым мечением терминальной дезоксинуклеотидилтрансферазы (TUNEL), фрагментацией ДНК и образованием апоптотических телец [145, 148]. САМ структурно аналогичны сфинганину и являются таким образом эффективные индукторы PCD.Возникающий механизм их действия заключается в том, что SAM могут конкурентно связываться с CerS в клетках. Такое связывание приводит к накоплению свободных сфингоидных оснований, субстратов CerS, в то время как церамиды как продукты CerS потребляются и восстанавливаются, активируя PCD в линиях клеток растений и животных [3,85]. Например, сообщалось, что FB 1 является мощным конкурентным ингибитором CerS из микросом печени и мозга в нескольких линиях клеток млекопитающих [149, 150].
Взаимосвязь между апоптозом и передачей сигналов церамидов была установлена как у растений, так и у животных в их ответе на SAM [120].Например, индукция гибели клеток как в клетках почек томатов, так и в клетках почки африканской зеленой мартышки (CV-1) происходила при одинаковых концентрациях токсина и временных рамках. Для обоих типов клеток наблюдались морфологические маркеры, характерные для апоптоза, включая клетки с положительным концевым мечением терминальной дезоксинуклеотидилтрансферазы (TUNEL), фрагментацией ДНК и образованием апоптотических телец [145, 148]. САМ структурно аналогичны сфинганину и являются таким образом эффективные индукторы PCD.Возникающий механизм их действия заключается в том, что SAM могут конкурентно связываться с CerS в клетках. Такое связывание приводит к накоплению свободных сфингоидных оснований, субстратов CerS, в то время как церамиды как продукты CerS потребляются и восстанавливаются, активируя PCD в линиях клеток растений и животных [3,85]. Например, сообщалось, что FB 1 является мощным конкурентным ингибитором CerS из микросом печени и мозга в нескольких линиях клеток млекопитающих [149, 150]. Увеличение сфинганина наблюдалось в тесте in vivo на ингибирование CerS, а также у животных, получавших FB, получавших высокие концентрации [2].Кроме того, TA и FB 1 могут ингибировать CerS в гепатоцитах крыс и плодах зеленых томатов [150, 151]. Кроме того, было обнаружено, что FB -1 не только индуцировал апоптоз в клетках животных, но также изменял морфологию клеток, межклеточные взаимодействия, поведение белков клеточной поверхности, активность протеинкиназ, а также рост и жизнеспособность клеток в неапоптотических клетках [148,152 ]. В клетках растений после воздействия SAM концентрация сфингозина значительно увеличивалась в течение короткого времени, после чего следовало накопление ROS в цитоплазме и затем апоптоз.Эти результаты свидетельствуют о том, что накопление сфингозина в клетках является сигналом АФК, ведущим к гибели клеток [43,44,85]. Индукция PCD под действием FB 1 также связана с накоплением церамидов. У A. thaliana есть два типа CerS, которые используют разные субстраты.
Увеличение сфинганина наблюдалось в тесте in vivo на ингибирование CerS, а также у животных, получавших FB, получавших высокие концентрации [2].Кроме того, TA и FB 1 могут ингибировать CerS в гепатоцитах крыс и плодах зеленых томатов [150, 151]. Кроме того, было обнаружено, что FB -1 не только индуцировал апоптоз в клетках животных, но также изменял морфологию клеток, межклеточные взаимодействия, поведение белков клеточной поверхности, активность протеинкиназ, а также рост и жизнеспособность клеток в неапоптотических клетках [148,152 ]. В клетках растений после воздействия SAM концентрация сфингозина значительно увеличивалась в течение короткого времени, после чего следовало накопление ROS в цитоплазме и затем апоптоз.Эти результаты свидетельствуют о том, что накопление сфингозина в клетках является сигналом АФК, ведущим к гибели клеток [43,44,85]. Индукция PCD под действием FB 1 также связана с накоплением церамидов. У A. thaliana есть два типа CerS, которые используют разные субстраты. CerS класса I используют сфинганин и C-16 жирный ацил-CoA в качестве субстратов, тогда как класс II использует фитосфингозин и очень длинноцепочечный жирный ацил-CoA в качестве субстратов. FB 1 в основном подавляет активность CerS класса II.При обработке FB 1 содержание фитосфингозина в клетках значительно увеличивается. В то же время, как и продукт предыдущего шага, количество сфинганина также увеличивается, что обеспечивает большее количество субстратов для CerS класса I. Следовательно, продукты CerS класса I в клетках увеличиваются, что приводит к индуцированной PCD [153,154].
CerS класса I используют сфинганин и C-16 жирный ацил-CoA в качестве субстратов, тогда как класс II использует фитосфингозин и очень длинноцепочечный жирный ацил-CoA в качестве субстратов. FB 1 в основном подавляет активность CerS класса II.При обработке FB 1 содержание фитосфингозина в клетках значительно увеличивается. В то же время, как и продукт предыдущего шага, количество сфинганина также увеличивается, что обеспечивает большее количество субстратов для CerS класса I. Следовательно, продукты CerS класса I в клетках увеличиваются, что приводит к индуцированной PCD [153,154].7. Устойчивость растений к SAM
Фитогормоны также участвуют в защитной реакции, вызванной SAM. Изменения в этилене (ЭТ) были впервые обнаружены при некрозе томатов, вызванном ААЛ [155].Изменение восприятия этилена у «незрелых» мутантов томатов может заметно облегчить повреждение тканей, вызванное SAM, что указывает на передачу сигнала, связанную с этиленом, во время гибели растительных клеток [156]. Позже был проведен транскрипционный анализ гибели клеток, вызванной AAL-токсином, у Arabidopsis. Гены, реагирующие на АФК и ЕТ, были одними из самых ранних генов с повышенной экспрессией [157]. Mase использовала VIGS (вирус-индуцированное молчание гена) и подтвердила, что путь передачи сигналов ЕТ и каскады MAPK необходимы для индуцированного токсином AAL PCD у табака [158].Посредством SA-опосредованной супрессии ET глутатион (GSH) может быть вовлечен в резистентность в первую очередь к стрессу, вызванному AAL-токсином, у Arabidopsis [159]. В отличие от этилена в базальных защитных ответах хозяина против патотипа томатов A. alternata, жасмоната (JA ) -зависимый сигнальный путь не участвует в защите хозяина от токсигенного патогена A. alternata. JA влияет на приемлемость патогенов посредством токсин-независимого взаимодействия между растениями и токсикогенными патогенами-некротрофами.Он может действовать до биосинтеза этилена в гибели клеток томата, вызванной AAL-токсином [160, 161]. Позже сравнительный протеомный анализ показал, что зависимый от COI1 (нечувствительный к коронатину 1, рецептор JA) путь JA усиливает ПКС, индуцированную ААЛ-токсином, посредством регулирования окислительно-восстановительного статуса листьев, других путей фитогормона и / или важных компонентов ПКС.
Позже был проведен транскрипционный анализ гибели клеток, вызванной AAL-токсином, у Arabidopsis. Гены, реагирующие на АФК и ЕТ, были одними из самых ранних генов с повышенной экспрессией [157]. Mase использовала VIGS (вирус-индуцированное молчание гена) и подтвердила, что путь передачи сигналов ЕТ и каскады MAPK необходимы для индуцированного токсином AAL PCD у табака [158].Посредством SA-опосредованной супрессии ET глутатион (GSH) может быть вовлечен в резистентность в первую очередь к стрессу, вызванному AAL-токсином, у Arabidopsis [159]. В отличие от этилена в базальных защитных ответах хозяина против патотипа томатов A. alternata, жасмоната (JA ) -зависимый сигнальный путь не участвует в защите хозяина от токсигенного патогена A. alternata. JA влияет на приемлемость патогенов посредством токсин-независимого взаимодействия между растениями и токсикогенными патогенами-некротрофами.Он может действовать до биосинтеза этилена в гибели клеток томата, вызванной AAL-токсином [160, 161]. Позже сравнительный протеомный анализ показал, что зависимый от COI1 (нечувствительный к коронатину 1, рецептор JA) путь JA усиливает ПКС, индуцированную ААЛ-токсином, посредством регулирования окислительно-восстановительного статуса листьев, других путей фитогормона и / или важных компонентов ПКС. [162]. Чувствительность растений томата к грибковому возбудителю A. alternata f. sp. lycopersici контролируется локусом устойчивости к раку ствола Alternaria (Asc-локус) на хромосоме 3 [163].Мутации локуса Asc томата придают устойчивость к патогену, в то время как сверхэкспрессия гена Asc-1 томата обеспечивает высокую нечувствительность к SAM в томатах и придает устойчивость к инфекции патогенами чувствительных растений Nicotiana [164, 165]. Asc-1 является гомологом гена обеспечения долголетия дрожжей LAG1, который кодирует компоненты сфинганин-N-ацилтрансферазы. Этот ген устойчивости может предотвратить нарушение метаболизма сфинголипидов во время PCD, индуцированного AAL-токсином. И роды Nicotiana, и Lycopersicon принадлежат к Solanaceae.У томатов нечувствительность к SAM и восприимчивость к патогену определяется Asc-1 [166]. Напротив, SAM-чувствительные виды у Nicotiana (за исключением N. umbratica) все еще имеют гомологи Asc-1 и устойчивы к A. alternata f. sp. lycopersici с HR, что указывает на дополнительный (не связанный с хозяином) механизм устойчивости между Nicotiana и этим патогеном [167].
[162]. Чувствительность растений томата к грибковому возбудителю A. alternata f. sp. lycopersici контролируется локусом устойчивости к раку ствола Alternaria (Asc-локус) на хромосоме 3 [163].Мутации локуса Asc томата придают устойчивость к патогену, в то время как сверхэкспрессия гена Asc-1 томата обеспечивает высокую нечувствительность к SAM в томатах и придает устойчивость к инфекции патогенами чувствительных растений Nicotiana [164, 165]. Asc-1 является гомологом гена обеспечения долголетия дрожжей LAG1, который кодирует компоненты сфинганин-N-ацилтрансферазы. Этот ген устойчивости может предотвратить нарушение метаболизма сфинголипидов во время PCD, индуцированного AAL-токсином. И роды Nicotiana, и Lycopersicon принадлежат к Solanaceae.У томатов нечувствительность к SAM и восприимчивость к патогену определяется Asc-1 [166]. Напротив, SAM-чувствительные виды у Nicotiana (за исключением N. umbratica) все еще имеют гомологи Asc-1 и устойчивы к A. alternata f. sp. lycopersici с HR, что указывает на дополнительный (не связанный с хозяином) механизм устойчивости между Nicotiana и этим патогеном [167]. Многослойные системы защиты также существуют у Arabidopsis, не являющегося хозяином устойчивости к A. alternata [168]. Точно так же, хотя многие виды Fusarium продуцируют фумонизин, они не могут инфицировать AAL-чувствительные томаты.Эта нечувствительность к хозяину включает в себя многослойную систему защиты, включающую как до, так и после инвазии, и помогает растениям защищаться от различных патогенов [169]. Кроме того, Зеликур продемонстрировал, что два из трех гомологов Lag1 в геноме Orobanche cumana ответственны за повышенную чувствительность к AAL-токсину [170]. Помимо вышеупомянутых генов, несколько других генов были идентифицированы у Arabidopsis и, как было установлено, участвуют в AAL-индуцированном пути PCD, включая цинк A. thaliana 11 (белок цинкового пальца ZAT11), fbr41 (FB 1 Resistant41) и ген бакуловируса p35 (ингибитор класса цистеиновых протеаз).Все они показали защитное действие в отношении гибели клеток, вызванной AAL-токсином, и инфицирования растений патогенами [171, 172, 173].
Многослойные системы защиты также существуют у Arabidopsis, не являющегося хозяином устойчивости к A. alternata [168]. Точно так же, хотя многие виды Fusarium продуцируют фумонизин, они не могут инфицировать AAL-чувствительные томаты.Эта нечувствительность к хозяину включает в себя многослойную систему защиты, включающую как до, так и после инвазии, и помогает растениям защищаться от различных патогенов [169]. Кроме того, Зеликур продемонстрировал, что два из трех гомологов Lag1 в геноме Orobanche cumana ответственны за повышенную чувствительность к AAL-токсину [170]. Помимо вышеупомянутых генов, несколько других генов были идентифицированы у Arabidopsis и, как было установлено, участвуют в AAL-индуцированном пути PCD, включая цинк A. thaliana 11 (белок цинкового пальца ZAT11), fbr41 (FB 1 Resistant41) и ген бакуловируса p35 (ингибитор класса цистеиновых протеаз).Все они показали защитное действие в отношении гибели клеток, вызванной AAL-токсином, и инфицирования растений патогенами [171, 172, 173]. Открытие устойчивых генов обеспечило потенциальную стратегию контроля SAM в растениеводстве с помощью трансгенной модификации растений.
Открытие устойчивых генов обеспечило потенциальную стратегию контроля SAM в растениеводстве с помощью трансгенной модификации растений.Из-за высокой токсичности фуминизинов, особенно FB 1 , им было уделено большое количество исследований. До сих пор механизм токсичности FB -1 был сосредоточен на его структурном сходстве со сфинганином и последующем конкурентном ингибировании CerS и нарушении липидомных профилей.Однако появляются новые доказательства того, что FB 1 может нарушать функцию митохондрий и генерировать чрезмерно токсичные АФК.
В таблице 2 представлен список обзоров, обобщающих последние достижения, связанные с фумонизинами, в их оценке, биосинтезе, обнаружении, селекции устойчивых сортов и токсичности.8. Гены, ответственные за продукцию SAM
У грибов гены, непосредственно участвующие в пути биосинтеза одного и того же вторичного метаболита, обычно расположены в соседних положениях в кластерах генов в геноме [191]. Эти гены часто коэкспрессируются и совместно регулируются. Гены, участвующие в биосинтезе фумонизина, соответствуют этой общей схеме. В частности, виды Fusarium, способные продуцировать фумонизины, обычно содержат один кластер генов, участвующий в их синтезе, называемый кластером генов FUM. В настоящее время в кластерах генов FUM различных видов идентифицирован в общей сложности 21 ген и подтверждено участие в биосинтезе и регуляции или самозащите фумонизина с использованием различных подходов, таких как нокаут генов, замена доменов и гетерологичная экспрессия ( Таблица 3) [192,193,194,195,196,197].У F. verticillioides кластер FUM, ответственный за биосинтез фумонизина B, включает 17 генов [193,197,198]. Как описано ранее, SAM представляют собой соединения, производные от поликетидов, со структурным сходством со сфинганином. Поликетиды синтезируются поликетидсинтазами (PKS), которые представляют собой большие многофункциональные ферменты. FUM1 — это ген PKS, ранее обозначенный как FUM5 [199].
Эти гены часто коэкспрессируются и совместно регулируются. Гены, участвующие в биосинтезе фумонизина, соответствуют этой общей схеме. В частности, виды Fusarium, способные продуцировать фумонизины, обычно содержат один кластер генов, участвующий в их синтезе, называемый кластером генов FUM. В настоящее время в кластерах генов FUM различных видов идентифицирован в общей сложности 21 ген и подтверждено участие в биосинтезе и регуляции или самозащите фумонизина с использованием различных подходов, таких как нокаут генов, замена доменов и гетерологичная экспрессия ( Таблица 3) [192,193,194,195,196,197].У F. verticillioides кластер FUM, ответственный за биосинтез фумонизина B, включает 17 генов [193,197,198]. Как описано ранее, SAM представляют собой соединения, производные от поликетидов, со структурным сходством со сфинганином. Поликетиды синтезируются поликетидсинтазами (PKS), которые представляют собой большие многофункциональные ферменты. FUM1 — это ген PKS, ранее обозначенный как FUM5 [199]. PKS, кодируемый FUM1, катализирует синтез предшественника октадекановой кислоты в качестве начальной стадии биосинтеза FB в Fusarium spp.[200,201]. Предлагаемый путь биосинтеза FB описан на Рисунке 4 [197]. Вышеупомянутый предшественник подвергается конденсации с L-аланином для синтеза основной цепи поликетида, эта реакция катализируется аминотрансферазой Fum8 [192,201]. У штамма F. oxysporum O-1890 ортолог FUM8 определяет, что Fusarium продуцирует преимущественно FC [195]. Считается, что делеция fum8 в некоторых пятнах A. welwitschiae связана с потерей продукции FB 2 [202].Вероятный митохондриальный белок-носитель, кодируемый FUM11, транспортирует трикарбоксилат субстрата для Fum7 (дегидрогеназа) и Fum10 (ацил-КоА-синтаза), чтобы продуцировать CoA-активированную трикарбаллиловую кислоту, которые присоединяются к основной цепи поликетида с помощью Fum14 (белок домена конденсации) [192,194 ]. Обеспечивающие этапы биосинтеза фумонизина, включающие различные модификации основной цепи (включая в первую очередь гидроксилирование), катализируются несколькими ферментами, такими как Fum6 / Fum12 / Fum15 (монооксигеназа цитохрома P450), Fum2 (гидроксилаза C10), Fum13 (короткоцепочечная дегидрогеназа / 3-кеторедукция) и, наконец, Fum3 (гидроксилаза C5, диоксигеназа), катализируемая белком, кодируемым FUM9 (аллели FUM3) [192,193,203,204,205,206].
PKS, кодируемый FUM1, катализирует синтез предшественника октадекановой кислоты в качестве начальной стадии биосинтеза FB в Fusarium spp.[200,201]. Предлагаемый путь биосинтеза FB описан на Рисунке 4 [197]. Вышеупомянутый предшественник подвергается конденсации с L-аланином для синтеза основной цепи поликетида, эта реакция катализируется аминотрансферазой Fum8 [192,201]. У штамма F. oxysporum O-1890 ортолог FUM8 определяет, что Fusarium продуцирует преимущественно FC [195]. Считается, что делеция fum8 в некоторых пятнах A. welwitschiae связана с потерей продукции FB 2 [202].Вероятный митохондриальный белок-носитель, кодируемый FUM11, транспортирует трикарбоксилат субстрата для Fum7 (дегидрогеназа) и Fum10 (ацил-КоА-синтаза), чтобы продуцировать CoA-активированную трикарбаллиловую кислоту, которые присоединяются к основной цепи поликетида с помощью Fum14 (белок домена конденсации) [192,194 ]. Обеспечивающие этапы биосинтеза фумонизина, включающие различные модификации основной цепи (включая в первую очередь гидроксилирование), катализируются несколькими ферментами, такими как Fum6 / Fum12 / Fum15 (монооксигеназа цитохрома P450), Fum2 (гидроксилаза C10), Fum13 (короткоцепочечная дегидрогеназа / 3-кеторедукция) и, наконец, Fum3 (гидроксилаза C5, диоксигеназа), катализируемая белком, кодируемым FUM9 (аллели FUM3) [192,193,203,204,205,206]. Кроме того, было подтверждено, что FUM21, кодирующий GAL4-подобный фактор транскрипции Zn (II) 2Cys6, участвует в регуляции синтеза фумонизина. Однако оказалось, что делеция FUM16 не оказала явного влияния на продукцию фумонизина у F. verticillioides [194,198,207,208,209]. Недавно было обнаружено, что FUM17 – FUM19 у F. verticillioides помогает грибку избежать собственной токсичности во время производства фумонизина. Fum19 представляет собой переносчик АТФ-связывающей кассеты (переносчик ABC) и действует как репрессор кластера генов FUM.FUM17 и FUM18 являются гомологами CerS. FUM18 может полностью комплементировать дрожжевой CerS-нулевой мутант LAG1 / LAC1, тогда как коэкспрессия FUM17 и CER3 частично дополняется. Белки Fum17 и Fum18 позволяют F. verticillioides повышать свою устойчивость к фумонизину, обеспечивая закодированную кластером FUM активность CerS в качестве первого уровня самозащиты [197]. Помимо генов FUM, другие гены, такие как FST1 (транспортер), FUG1 (факторы транскрипции или передачи сигнала), CPP1 (каталитическая субъединица протеинфосфатазы типа 2A) и FvVEl (регулятор) в F.
Кроме того, было подтверждено, что FUM21, кодирующий GAL4-подобный фактор транскрипции Zn (II) 2Cys6, участвует в регуляции синтеза фумонизина. Однако оказалось, что делеция FUM16 не оказала явного влияния на продукцию фумонизина у F. verticillioides [194,198,207,208,209]. Недавно было обнаружено, что FUM17 – FUM19 у F. verticillioides помогает грибку избежать собственной токсичности во время производства фумонизина. Fum19 представляет собой переносчик АТФ-связывающей кассеты (переносчик ABC) и действует как репрессор кластера генов FUM.FUM17 и FUM18 являются гомологами CerS. FUM18 может полностью комплементировать дрожжевой CerS-нулевой мутант LAG1 / LAC1, тогда как коэкспрессия FUM17 и CER3 частично дополняется. Белки Fum17 и Fum18 позволяют F. verticillioides повышать свою устойчивость к фумонизину, обеспечивая закодированную кластером FUM активность CerS в качестве первого уровня самозащиты [197]. Помимо генов FUM, другие гены, такие как FST1 (транспортер), FUG1 (факторы транскрипции или передачи сигнала), CPP1 (каталитическая субъединица протеинфосфатазы типа 2A) и FvVEl (регулятор) в F. verticillioides, PKS3 и PKS11 у F. proliferatum и факторы транскрипции GATA-типа AreA и AreB (известные как глобальные регуляторы азота) у F. fujikuroi также, как было показано, играют важную роль в биосинтезе и регуляции фумонизина [209, 210, 211, 212, 213, 214]. Кроме того, вырожденный, чрезмерно представленный мотив, который потенциально участвует в цис-регуляции генов FUM и биосинтезе фумонизина, также был идентифицирован как у F. verticillioides, так и у Asp. niger, но не обнаружен у грибов, не продуцирующих фумонизин, содержащих различные гомологи ФУМ [215].Было обнаружено, что несколько абиотических и биотических факторов влияют на экспрессию генов FUM и регулируют биосинтез фумонизина. Эти факторы включают активность воды, температуру, источники углерода и других питательных веществ, виды и разновидности растений-хозяев или их экстракты, а также возраст растений [216 217 218 219 220]. Зрелые растения и экстракты этих растений часто связаны с более высокими концентрациями SAM. Было высказано предположение, что сбор урожая на более ранних стадиях, кроме полной зрелости, мог бы быть одной из стратегий борьбы с загрязнением фумонизином [221, 222].
verticillioides, PKS3 и PKS11 у F. proliferatum и факторы транскрипции GATA-типа AreA и AreB (известные как глобальные регуляторы азота) у F. fujikuroi также, как было показано, играют важную роль в биосинтезе и регуляции фумонизина [209, 210, 211, 212, 213, 214]. Кроме того, вырожденный, чрезмерно представленный мотив, который потенциально участвует в цис-регуляции генов FUM и биосинтезе фумонизина, также был идентифицирован как у F. verticillioides, так и у Asp. niger, но не обнаружен у грибов, не продуцирующих фумонизин, содержащих различные гомологи ФУМ [215].Было обнаружено, что несколько абиотических и биотических факторов влияют на экспрессию генов FUM и регулируют биосинтез фумонизина. Эти факторы включают активность воды, температуру, источники углерода и других питательных веществ, виды и разновидности растений-хозяев или их экстракты, а также возраст растений [216 217 218 219 220]. Зрелые растения и экстракты этих растений часто связаны с более высокими концентрациями SAM. Было высказано предположение, что сбор урожая на более ранних стадиях, кроме полной зрелости, мог бы быть одной из стратегий борьбы с загрязнением фумонизином [221, 222]. Секвенирование и анализ генома Asp. niger показал, что его геном содержит кластер генов (кластер fum), гомологичный кластеру FUM у видов Fusarium (показано в таблице 3). В частности, было обнаружено 12 гомологов генов синтеза фумонизина, включая гены fum1, fum3, fum6, fum7, fum8, fum10, fum13 и fum16, fum19 и fum21 [208, 223, 224]. Этот кластер генов также обнаружен в изолятах Asp, продуцирующих фумонизин. welwitschiae, но отсутствует в геномах других секвенированных Aspergilli, не продуцирующих фумонизин, таких как Asp.fumigatus, Asp. oryzae и Asp. нидуланы [7,208]. Кроме того, гомологи нескольких генов fum были обнаружены у нескольких других Aspergilli spp. но там, где производство фумонизина не было обнаружено (сведено в Таблицу 4). Некоторые виды Aspergullus spp. содержат гены, которые уникальны для них. Например, ген дегидрогеназы (sdr1) с короткой длиной цепи был обнаружен в кластере fum Asp. niger, но отсутствует в кластере генов FUM Fusarium spp. Напротив, ген Fusarium FUM2 с функцией гидроксилирования в положении C-10 основной цепи фумонизина отсутствует в Asp.
Секвенирование и анализ генома Asp. niger показал, что его геном содержит кластер генов (кластер fum), гомологичный кластеру FUM у видов Fusarium (показано в таблице 3). В частности, было обнаружено 12 гомологов генов синтеза фумонизина, включая гены fum1, fum3, fum6, fum7, fum8, fum10, fum13 и fum16, fum19 и fum21 [208, 223, 224]. Этот кластер генов также обнаружен в изолятах Asp, продуцирующих фумонизин. welwitschiae, но отсутствует в геномах других секвенированных Aspergilli, не продуцирующих фумонизин, таких как Asp.fumigatus, Asp. oryzae и Asp. нидуланы [7,208]. Кроме того, гомологи нескольких генов fum были обнаружены у нескольких других Aspergilli spp. но там, где производство фумонизина не было обнаружено (сведено в Таблицу 4). Некоторые виды Aspergullus spp. содержат гены, которые уникальны для них. Например, ген дегидрогеназы (sdr1) с короткой длиной цепи был обнаружен в кластере fum Asp. niger, но отсутствует в кластере генов FUM Fusarium spp. Напротив, ген Fusarium FUM2 с функцией гидроксилирования в положении C-10 основной цепи фумонизина отсутствует в Asp. niger fum cluster [207, 208]. Этот результат согласуется с исследованием, показывающим, что Asp. niger производит только фумонизины FB 2 , FB 4 и FB 6 , в которых отсутствует гидроксил на C-10 [7,225,226]. Однако изоляты нескольких черных аспергилл (включая A. niger, Asp. Foetidus и A. tubingensis), выделенные из арахиса и кукурузы, также продуцировали FB 1 и FB 3 , что согласуется со сложной картиной биосинтеза фумонизинов в Aspergilli виды [227].
niger fum cluster [207, 208]. Этот результат согласуется с исследованием, показывающим, что Asp. niger производит только фумонизины FB 2 , FB 4 и FB 6 , в которых отсутствует гидроксил на C-10 [7,225,226]. Однако изоляты нескольких черных аспергилл (включая A. niger, Asp. Foetidus и A. tubingensis), выделенные из арахиса и кукурузы, также продуцировали FB 1 и FB 3 , что согласуется со сложной картиной биосинтеза фумонизинов в Aspergilli виды [227].Таблица 3. Гомологичные гены и их функциональные роли в биосинтезе SAM.
Таблица 3. Гомологичные гены и их функциональные роли в биосинтезе SAM.
| Гомолог кластерных генов | Предсказать продукт и функцию гена | Ссылка | |||||||
|---|---|---|---|---|---|---|---|---|---|
| фумонизин | AAL-токсин | Aspergillus spp. | A. alternata | [169,224] | |||||
| FUM1 (ген 1 биосинтеза фумонизина, ранее обозначенный как FUM5)) | fum1 | ALT1 (ген AAL-синтетический токсин ) 9176 | [12,199,200] | ||||||
| FUM2 | отсутствует | Диоксигеназа для гидроксилирования C10 | [228] | ||||||
| FUM3 | fum6 | гидрокислорода Fum6 )[205,228,229] | |||||||
| FUM4 | Не ясно | [230] | |||||||
| FUM6 | fum6 | Циктохромная оксигеназа C1550 CYT2 | [192,231] | ||||||
| FUM7 | fum7 | ALT3 | Спирт типа III de гидрогеназы для образования боковой цепи PTCA (пропан-1,2,3-трикарбоновая кислота) | [192,194,232] | |||||
| FUM8 | fum8 | α-оксоаминсинтаза и гомологична для переноса аминогруппы и образования | / FC[192,195] | ||||||
| FUM10 | fum10 | Ацил-КоА-жирная синтаза для этерификации PTCA | [193,194] | ||||||
| FUM11 | транспорт митохондриального белка FUM11 | 9 ] | |||||||
| FUM12 | монооксигеназы цитохрома P450 | [193] | |||||||
| FUM13 | fum13 | ALT6,203 | кетодегидроген | Short-of-chain FUM14 | fum14 | Нерибосомальная пептидная синтетаза для этерификации PTCA ион | [193,194,233] | ||
| FUM15 | fum15 | Монооксигеназы цитохрома P450 | [193] | ||||||
| FUM16 | FUM16 | fum6 | |||||||
| FUM17 | CerS для самозащиты от фумонизинов | [193,197] | |||||||
| FUM18 | CerS для самозащиты от фумонизинов 9193 | 9194,1 | fum19 | Транспортный белок ABC как репрессор кластера генов FUM | [193,197] | ||||
| FUM20 | Неясно | [234] | [234] | Фактор транскрипции Zn (II) 2Cys6 | [198] | 9038 3отсутствует | SDR1 | Короткоцепочечная дегидрогеназа / редуктаза (SDR) | [7] |
 Fusar3.
Fusar3. alternata f. sp. Lycopersici, специфический патотип распространенного грибкового патогена растений в роду, отличном от Aspergillus и Fusarium. Этот патотип может продуцировать производные поликетида, сходные по структуре с фумонизинами, продуцируемыми видами Fusarium. Гены ALT (синтез AAL-токсинов) также располагаются в виде кластера на условной одноразовой (CD) хромосоме размером ~ 1.0 Mb у всех штаммов томатного патотипа A. alternata из разных стран [237]. Такие CD-хромосомы, несущие кластер генов биосинтеза токсинов, были обнаружены и у других патотипов A.alternata [169]. Они контролируют продукцию других HST и их патогенность для хозяина. Они могли стабильно сохраняться на новом генетическом фоне с расширенным диапазоном патогенности, что было подтверждено тестом слияния протопластов [238]. Кластер генов токсина AAL включает по меньшей мере 13 генов в области 120 т.п.н., некоторые из которых показали значительное сходство с кластером генов FUM, состоящим из 17 генов в области 45,5 т.
alternata f. sp. Lycopersici, специфический патотип распространенного грибкового патогена растений в роду, отличном от Aspergillus и Fusarium. Этот патотип может продуцировать производные поликетида, сходные по структуре с фумонизинами, продуцируемыми видами Fusarium. Гены ALT (синтез AAL-токсинов) также располагаются в виде кластера на условной одноразовой (CD) хромосоме размером ~ 1.0 Mb у всех штаммов томатного патотипа A. alternata из разных стран [237]. Такие CD-хромосомы, несущие кластер генов биосинтеза токсинов, были обнаружены и у других патотипов A.alternata [169]. Они контролируют продукцию других HST и их патогенность для хозяина. Они могли стабильно сохраняться на новом генетическом фоне с расширенным диапазоном патогенности, что было подтверждено тестом слияния протопластов [238]. Кластер генов токсина AAL включает по меньшей мере 13 генов в области 120 т.п.н., некоторые из которых показали значительное сходство с кластером генов FUM, состоящим из 17 генов в области 45,5 т. п.н. Однако расположение генов в кластерах ALT и FUM отличается у этих двух групп грибов.Кроме того, в одном штамме, As-27 A. alternata, на хромосоме CD присутствовало два набора кластера генов биосинтеза AAL-токсина [169]. Было обнаружено, что синтез AAL-токсинов инициируется ALT1-кодируемыми PKS с образованием остова аминопентола, который затем был модифицирован другими ферментами [239]. Функциональное сходство между ALT1 и FUM1 было подтверждено, когда биосинтез фумонизина в FUM1-разрушенном F. verticillioides был восстановлен при дополнении ALT1 из A. alternata [240]. Аналогичным образом, экспрессия ALT1 и продукция AAL-токсинов также регулируются глобальным регулятором LaeA [241].Накоплению AAL-токсина также способствует высокая активность воды (0,995 Вт) и высокая температура (выше 30 ° C) во время инкубационного периода патогена [242]. Интересно, что ген PKS, подобный FUM1, и ортологи кластера генов FUM были обнаружен в геноме Cochliobolus spp. филогенетическим анализом грибного поликетида.
п.н. Однако расположение генов в кластерах ALT и FUM отличается у этих двух групп грибов.Кроме того, в одном штамме, As-27 A. alternata, на хромосоме CD присутствовало два набора кластера генов биосинтеза AAL-токсина [169]. Было обнаружено, что синтез AAL-токсинов инициируется ALT1-кодируемыми PKS с образованием остова аминопентола, который затем был модифицирован другими ферментами [239]. Функциональное сходство между ALT1 и FUM1 было подтверждено, когда биосинтез фумонизина в FUM1-разрушенном F. verticillioides был восстановлен при дополнении ALT1 из A. alternata [240]. Аналогичным образом, экспрессия ALT1 и продукция AAL-токсинов также регулируются глобальным регулятором LaeA [241].Накоплению AAL-токсина также способствует высокая активность воды (0,995 Вт) и высокая температура (выше 30 ° C) во время инкубационного периода патогена [242]. Интересно, что ген PKS, подобный FUM1, и ортологи кластера генов FUM были обнаружен в геноме Cochliobolus spp. филогенетическим анализом грибного поликетида. Предполагалось, что эти грибы продуцируют фумонизин или другие SAM [243,244]. Чтобы предсказать возможное распределение продукции SAM в грибах, Kim et al. предложили гипотезу о кластерах биосинтетических генов SAM на основе модели биосинтеза фумонизина.Этот предполагаемый кластер генов должен включать PKS, аминотрансферазу и ген дегидрогеназы. Их модель показала, что шестьдесят девять видов рода Fusarium и виды двадцати четырех других родов грибов, по прогнозам, имеют по крайней мере один кластер SAM [245].
Предполагалось, что эти грибы продуцируют фумонизин или другие SAM [243,244]. Чтобы предсказать возможное распределение продукции SAM в грибах, Kim et al. предложили гипотезу о кластерах биосинтетических генов SAM на основе модели биосинтеза фумонизина.Этот предполагаемый кластер генов должен включать PKS, аминотрансферазу и ген дегидрогеназы. Их модель показала, что шестьдесят девять видов рода Fusarium и виды двадцати четырех других родов грибов, по прогнозам, имеют по крайней мере один кластер SAM [245].9. Эволюция продуцирования SAM
Горизонтальный перенос генов (HGT) был предложен в качестве основного механизма, ответственного за приобретение и эволюцию кластеров генов биосинтеза фумонизинов и AAL-токсинов среди дивергентных грибов [7,9,169,224].У Fusarium анализ последовательности генома показал, что гены биосинтеза фумонизина (FUM) сгруппированы и демонстрируют последовательную генную организацию у большинства видов. Например, кластеры FUM в F. oxysporum, F. proliferatum и F. verticillioides демонстрируют относительно небольшую изменчивость, при этом порядок и ориентация генов в кластерах одинаковы друг с другом. Кроме того, вариабельность их последовательностей среди ортологов кодирующих областей F. oxysporum и F. verticillioides относительно невелика [193,195].Два разных вида Aspergillus, которые продуцируют фумонизины, Asp. нигер и жерех. welwitschiae, также похожи друг на друга по порядку генов, но отличаются от кластера FUM у Fusarium [224]. В настоящее время томатный патотип A. alternata является единственным видом рода Alternaria, способным продуцировать SAM, и этот патотип имеет кластерные гены (участвующие в биосинтезе AAL-токсина), аналогичные кластеру FUM в Fusarium [169, 237]. В совокупности анализ структуры гена и последовательности показал, что кластер биосинтетических генов SAM, вероятно, возник в Fusarium и передан Asp.niger и A. alternata, разработанные HGT. Сходство химической структуры и цитотоксичности фумонизинов и AAL-токсинов для растений и животных также подтверждает эту гипотезу.
proliferatum и F. verticillioides демонстрируют относительно небольшую изменчивость, при этом порядок и ориентация генов в кластерах одинаковы друг с другом. Кроме того, вариабельность их последовательностей среди ортологов кодирующих областей F. oxysporum и F. verticillioides относительно невелика [193,195].Два разных вида Aspergillus, которые продуцируют фумонизины, Asp. нигер и жерех. welwitschiae, также похожи друг на друга по порядку генов, но отличаются от кластера FUM у Fusarium [224]. В настоящее время томатный патотип A. alternata является единственным видом рода Alternaria, способным продуцировать SAM, и этот патотип имеет кластерные гены (участвующие в биосинтезе AAL-токсина), аналогичные кластеру FUM в Fusarium [169, 237]. В совокупности анализ структуры гена и последовательности показал, что кластер биосинтетических генов SAM, вероятно, возник в Fusarium и передан Asp.niger и A. alternata, разработанные HGT. Сходство химической структуры и цитотоксичности фумонизинов и AAL-токсинов для растений и животных также подтверждает эту гипотезу. Тем не менее, различия между кластерами FUM и кластером генов биосинтеза AAL-токсинов также свидетельствуют о значительном расхождении между ними. Анализ кластера генов FUM среди видов Fusarium также выявил доказательства увеличения, потери и мутаций генов в различных генах. Например, не все виды Fusarium могут продуцировать фумонизины.Даже для видов, которые могут синтезировать фумонизины, некоторые штаммы производят больше, чем другие в тех же экспериментальных условиях, в то время как другие штаммы вообще не производят токсины [9,196]. Действительно, для некоторых штаммов, хотя гены FUM были обнаружены, фумонизин не был обнаружен. Вероятно, это было связано с накоплением мутаций, приводящих к нефункциональности генов FUM. Например, несколько мутаций были обнаружены в FUM7 и FUM21 у F. fujikuroi [246]. Кроме того, у некоторых видов Fusarium, не продуцирующих фумонизин, отсутствуют гены биосинтеза фумонизина, но сохранены гомологи нескольких генов, фланкирующих кластер Fum у F.verticillioides [247].
Тем не менее, различия между кластерами FUM и кластером генов биосинтеза AAL-токсинов также свидетельствуют о значительном расхождении между ними. Анализ кластера генов FUM среди видов Fusarium также выявил доказательства увеличения, потери и мутаций генов в различных генах. Например, не все виды Fusarium могут продуцировать фумонизины.Даже для видов, которые могут синтезировать фумонизины, некоторые штаммы производят больше, чем другие в тех же экспериментальных условиях, в то время как другие штаммы вообще не производят токсины [9,196]. Действительно, для некоторых штаммов, хотя гены FUM были обнаружены, фумонизин не был обнаружен. Вероятно, это было связано с накоплением мутаций, приводящих к нефункциональности генов FUM. Например, несколько мутаций были обнаружены в FUM7 и FUM21 у F. fujikuroi [246]. Кроме того, у некоторых видов Fusarium, не продуцирующих фумонизин, отсутствуют гены биосинтеза фумонизина, но сохранены гомологи нескольких генов, фланкирующих кластер Fum у F.verticillioides [247]. Интересно, что фланкирующие области кластера FUM часто различаются между видами, что согласуется с независимым происхождением кластера FUM, включая независимое приобретение и / или потерю кластера генов видами, продуцирующими фумонизин [195]. Например, не продуцирующие фумонизин штаммы F. verticillioides, выделенные из банана, не содержали кластера функционального гена биосинтеза фумонизина (FUM), но содержали части FUM21 и FUM19, фланкирующие кластер, оба из которых являются концевыми генами на каждом конце. кластера ФУМ.Тем не менее, штаммы бананов по-прежнему патогенны для бананов, но они не проявляют той же патологии, как штаммы, продуцирующие фумонизин, для кукурузы. Когда штамм банана был котрансформирован двумя перекрывающимися космидами, содержащими весь кластер генов FUM, продукция фумонизина и патогенность проростков кукурузы были восстановлены [248]. Подобно Fusarium, FB-непродуцирующие изоляты Asp. niger или другие виды Aspergillus также имели интактный кластер fum или несколько паттернов делеции гена fum, соответственно (показано в таблице 4).
Интересно, что фланкирующие области кластера FUM часто различаются между видами, что согласуется с независимым происхождением кластера FUM, включая независимое приобретение и / или потерю кластера генов видами, продуцирующими фумонизин [195]. Например, не продуцирующие фумонизин штаммы F. verticillioides, выделенные из банана, не содержали кластера функционального гена биосинтеза фумонизина (FUM), но содержали части FUM21 и FUM19, фланкирующие кластер, оба из которых являются концевыми генами на каждом конце. кластера ФУМ.Тем не менее, штаммы бананов по-прежнему патогенны для бананов, но они не проявляют той же патологии, как штаммы, продуцирующие фумонизин, для кукурузы. Когда штамм банана был котрансформирован двумя перекрывающимися космидами, содержащими весь кластер генов FUM, продукция фумонизина и патогенность проростков кукурузы были восстановлены [248]. Подобно Fusarium, FB-непродуцирующие изоляты Asp. niger или другие виды Aspergillus также имели интактный кластер fum или несколько паттернов делеции гена fum, соответственно (показано в таблице 4). Точно так же кластер генов, продуцирующих AAL-токсин у A. alternata, вероятно, произошел от видов Fusarium в результате HGT-события. Данные кластера генов ALT, распределенного в изолятах A. alternata, также подтверждают эту гипотезу о HGT в патогенах, продуцирующих AAL-токсин [237]. Вместе эти результаты предполагают, что было несколько HGT кластера между видами, а также дублирование и потеря всего или части кластера после приобретения [9]. Гипотеза о том, что несколько HGT были вовлечены в создание текущего распределения Кластеры генов FUM также подтверждаются филогенетическими исследованиями.В частности, филогенетические деревья, основанные на генах из кластера FUM, часто не параллельны дереву видов Fusarium, основанному на других генах [9]. У Fusarium ген фактора удлинения трансляции (tef-1α) является наиболее часто используемым геном-маркером для таксономических исследований. Однако последовательности tef-1α часто недостаточны для различения изолятов, продуцирующих фумонизин, из разных стран и / или растений-хозяев.
Точно так же кластер генов, продуцирующих AAL-токсин у A. alternata, вероятно, произошел от видов Fusarium в результате HGT-события. Данные кластера генов ALT, распределенного в изолятах A. alternata, также подтверждают эту гипотезу о HGT в патогенах, продуцирующих AAL-токсин [237]. Вместе эти результаты предполагают, что было несколько HGT кластера между видами, а также дублирование и потеря всего или части кластера после приобретения [9]. Гипотеза о том, что несколько HGT были вовлечены в создание текущего распределения Кластеры генов FUM также подтверждаются филогенетическими исследованиями.В частности, филогенетические деревья, основанные на генах из кластера FUM, часто не параллельны дереву видов Fusarium, основанному на других генах [9]. У Fusarium ген фактора удлинения трансляции (tef-1α) является наиболее часто используемым геном-маркером для таксономических исследований. Однако последовательности tef-1α часто недостаточны для различения изолятов, продуцирующих фумонизин, из разных стран и / или растений-хозяев. Напротив, полиморфизмы последовательностей ДНК на основе FUM1 часто обеспечивают лучшее разрешение среди патотипов [196,249,250,251,252,253].Например, филогенетический анализ 38 изолятов F. proliferatum, происходящих от разных хозяев, показал, что вариабельность последовательностей среди штаммов в гене FUM1 коррелировала с вариацией последовательностей растений-хозяев. В частности, филогенетический анализ, основанный на частичных последовательностях FUM1, более четко дифференцировал группы, связанные с хозяином, чем анализ на основе последовательностей tef-1α. Лучшая выделенная группа состоит из изолятов, полученных из чеснока, и сформировала отдельную ветвь на дендрограмме на основе FUM1 [196, 254]. Аналогичным образом, анализ дивергенции последовательности FUM1 на F.proliferatum и F. verticillioides, выделенные из гороха, также составляли отдельную группу по сравнению со штаммами, полученными от разных видов хозяев [253]. Помимо различий генов между изолятами от разных растений-хозяев, вариации как токсигенного потенциала, так и характера роста могут также различаться между изоляты, полученные от одних и тех же растений-хозяев.
Напротив, полиморфизмы последовательностей ДНК на основе FUM1 часто обеспечивают лучшее разрешение среди патотипов [196,249,250,251,252,253].Например, филогенетический анализ 38 изолятов F. proliferatum, происходящих от разных хозяев, показал, что вариабельность последовательностей среди штаммов в гене FUM1 коррелировала с вариацией последовательностей растений-хозяев. В частности, филогенетический анализ, основанный на частичных последовательностях FUM1, более четко дифференцировал группы, связанные с хозяином, чем анализ на основе последовательностей tef-1α. Лучшая выделенная группа состоит из изолятов, полученных из чеснока, и сформировала отдельную ветвь на дендрограмме на основе FUM1 [196, 254]. Аналогичным образом, анализ дивергенции последовательности FUM1 на F.proliferatum и F. verticillioides, выделенные из гороха, также составляли отдельную группу по сравнению со штаммами, полученными от разных видов хозяев [253]. Помимо различий генов между изолятами от разных растений-хозяев, вариации как токсигенного потенциала, так и характера роста могут также различаться между изоляты, полученные от одних и тех же растений-хозяев. Хотя не наблюдалось различий в уровнях FB, измеренных среди семян гороха, продукция FB различалась между выбранными штаммами F. proliferatum в культурах риса [253].Среди всех этих тестовых изолятов F. proliferatum наиболее разнообразной группой изолятов, обнаруженных на данный момент, были изоляты кукурузы [196, 254]. Как межвидовые, так и внутривидовые вариации в уровне синтеза FB могут, по крайней мере, частично объясняться различиями последовательностей внутри FUM кластера.
Хотя не наблюдалось различий в уровнях FB, измеренных среди семян гороха, продукция FB различалась между выбранными штаммами F. proliferatum в культурах риса [253].Среди всех этих тестовых изолятов F. proliferatum наиболее разнообразной группой изолятов, обнаруженных на данный момент, были изоляты кукурузы [196, 254]. Как межвидовые, так и внутривидовые вариации в уровне синтеза FB могут, по крайней мере, частично объясняться различиями последовательностей внутри FUM кластера. Таким образом, проведенный анализ показал, что кластер генов FUM отвечает за биосинтез фумонизина. Мутация и делеция некоторых или всех генов в кластере могут привести к ограниченному производству фумонизина или его отсутствию, что приведет к более слабому развитию болезни патогена на растении-хозяине.Анализ последовательности показал, что A. alternata, вероятно, приобрела способность продуцировать AAL-токсин из-за HGT кластера генов SAM из видов Fusarium, продуцирующих фумонизин, с последующей независимой эволюцией во взаимодействии патоген-хозяин. Дивергентные паттерны дивергенции последовательностей генов биосинтеза токсинов могут объяснить различия между фумонизинами и AAL-токсинами как в их продукции, так и в их влиянии на взаимодействия хозяин-патоген.
Дивергентные паттерны дивергенции последовательностей генов биосинтеза токсинов могут объяснить различия между фумонизинами и AAL-токсинами как в их продукции, так и в их влиянии на взаимодействия хозяин-патоген.
10. Метод обнаружения SAM
Для обнаружения SAM были разработаны различные методы, включая ВЭЖХ с детектированием флуоресцентного / испарительного светорассеяния или масс-спектрометрию (МС), тонкослойную хроматографию (ТСХ), ELISA, преобразование Фурье в ближней инфракрасной области. (FT-NIR) спектроскопия и т.д. [255,256,257,258,259,260,261,262,263].Хотя эти традиционные аналитические методы были разработаны для обнаружения и количественной оценки известных соединений, для которых доступны стандарты, есть четкие доказательства того, что многие неизвестные производные могут присутствовать в пищевых продуктах и пищевых продуктах, и некоторые из них могут быть токсичными для животных, включая человека [39]. В 2015 году был разработан полуцелевой метод, сочетающий фильтрацию продуктовых ионов и быстрое переключение полярности для быстрого обнаружения всех известных фумонизинов и AAL-токсинов. Этим методом были также открыты некоторые новые структурно родственные возникающие токсины [26].Помимо разработки методов, нацеленных на потенциально новые SAM, о которых ранее не сообщалось, существуют также разработки эффективных методов, нацеленных на обнаружение известных SAM. Действительно, было разработано несколько новых методов, основанных на иммуноанализе, для простого, быстрого и сверхчувствительного количественного определения SAM на месте. Например, в одном методе использовались хемилюминесцентные биосенсоры, объединяющие иммуноанализ конкурентного бокового потока и камеру устройства с зарядовой связью для обнаружения FB [264, 265]. Другой метод использует наночастицы золота или наночастицы квантовых точек на основе моноклональных антител против фумонизина и позволяет быстро обнаружить этот микотоксин за один этап [266, 267, 268].Кроме того, анализ изотермической амплификации, опосредованной петлей (LAMP), основанный на обнаружении fum10, может специфически обнаружить гены, участвующие в биосинтезе FB 2 у видов Aspergillus, которые можно оценить невооруженным глазом за короткое время [269] .
Этим методом были также открыты некоторые новые структурно родственные возникающие токсины [26].Помимо разработки методов, нацеленных на потенциально новые SAM, о которых ранее не сообщалось, существуют также разработки эффективных методов, нацеленных на обнаружение известных SAM. Действительно, было разработано несколько новых методов, основанных на иммуноанализе, для простого, быстрого и сверхчувствительного количественного определения SAM на месте. Например, в одном методе использовались хемилюминесцентные биосенсоры, объединяющие иммуноанализ конкурентного бокового потока и камеру устройства с зарядовой связью для обнаружения FB [264, 265]. Другой метод использует наночастицы золота или наночастицы квантовых точек на основе моноклональных антител против фумонизина и позволяет быстро обнаружить этот микотоксин за один этап [266, 267, 268].Кроме того, анализ изотермической амплификации, опосредованной петлей (LAMP), основанный на обнаружении fum10, может специфически обнаружить гены, участвующие в биосинтезе FB 2 у видов Aspergillus, которые можно оценить невооруженным глазом за короткое время [269] . Этот метод также применялся для обнаружения FB 1 , нацеленного на ген FUM1 в Fusarium [270]. Прямое обнаружение генов, участвующих в биосинтезе SAM в системах сельскохозяйственного производства, позволяет проводить широкие оценки потенциальных штаммов, продуцирующих фумонизин, в пищевых продуктах и кормах.С этой целью мультиплексная ПЦР показала большие перспективы для обнаружения нескольких видов Fusarium и Aspergillus, продуцирующих фумонизин [271, 272, 273]. Хотя в настоящее время основное внимание уделяется известным SAM с известной токсичностью, появляется все больше свидетельств того, что некоторые из этих токсинов замаскированы и замаскированы. нелегко обнаружить или количественно оценить. Для обеспечения безопасности пищевых продуктов необходимо обнаруживать и количественно определять как свободные, так и замаскированные формы микотоксинов. Замаскированные микотоксины обычно представляют собой модифицированные формы микотоксинов растительными ферментами во время инфекции и обычно не обнаруживаются при обычном анализе.
Этот метод также применялся для обнаружения FB 1 , нацеленного на ген FUM1 в Fusarium [270]. Прямое обнаружение генов, участвующих в биосинтезе SAM в системах сельскохозяйственного производства, позволяет проводить широкие оценки потенциальных штаммов, продуцирующих фумонизин, в пищевых продуктах и кормах.С этой целью мультиплексная ПЦР показала большие перспективы для обнаружения нескольких видов Fusarium и Aspergillus, продуцирующих фумонизин [271, 272, 273]. Хотя в настоящее время основное внимание уделяется известным SAM с известной токсичностью, появляется все больше свидетельств того, что некоторые из этих токсинов замаскированы и замаскированы. нелегко обнаружить или количественно оценить. Для обеспечения безопасности пищевых продуктов необходимо обнаруживать и количественно определять как свободные, так и замаскированные формы микотоксинов. Замаскированные микотоксины обычно представляют собой модифицированные формы микотоксинов растительными ферментами во время инфекции и обычно не обнаруживаются при обычном анализе. Например, замаскированные микотоксины могут конъюгировать с полярными веществами, храниться в вакуоли в растворимой форме или связываться с макромолекулами и, таким образом, изменять свои физиологические свойства. Хотя замаскированные микотоксины часто менее токсичны, чем немаскированные формы, они могут легко превращаться в немаскированные формы токсинов, в том числе во время переваривания пищи [274, 275]. Наиболее типичными замаскированными микотоксинами являются модифицированные формы зеараленона (ZEN), DON и фумонизинов [276, 277, 278, 279]. Так называемые скрытые фумонизины могут образовывать нековалентные связи с пищевыми макрокомпонентами, такими как продукты на основе крахмала.В определенных ситуациях замаскированные фумонизины могут присутствовать в пище в количествах, намного превышающих количество свободных форм. Многие факторы могут влиять на относительные доли SAM в замаскированных формах, включая условия роста сельскохозяйственных культур, а также методы хранения и обработки пищевых продуктов [279,280,281,282].
Например, замаскированные микотоксины могут конъюгировать с полярными веществами, храниться в вакуоли в растворимой форме или связываться с макромолекулами и, таким образом, изменять свои физиологические свойства. Хотя замаскированные микотоксины часто менее токсичны, чем немаскированные формы, они могут легко превращаться в немаскированные формы токсинов, в том числе во время переваривания пищи [274, 275]. Наиболее типичными замаскированными микотоксинами являются модифицированные формы зеараленона (ZEN), DON и фумонизинов [276, 277, 278, 279]. Так называемые скрытые фумонизины могут образовывать нековалентные связи с пищевыми макрокомпонентами, такими как продукты на основе крахмала.В определенных ситуациях замаскированные фумонизины могут присутствовать в пище в количествах, намного превышающих количество свободных форм. Многие факторы могут влиять на относительные доли SAM в замаскированных формах, включая условия роста сельскохозяйственных культур, а также методы хранения и обработки пищевых продуктов [279,280,281,282]. Эти скрытые опасности требуют разработки новых методов, позволяющих обнаруживать и количественно определять замаскированные формы SAM, а также другие микотоксины.
Эти скрытые опасности требуют разработки новых методов, позволяющих обнаруживать и количественно определять замаскированные формы SAM, а также другие микотоксины.Открытие и проверка сигнатуры сывороточного протеина с шестью маркерами для диагностики активного туберкулеза легких
РЕЗЮМЕ
Новые тесты на биомаркеры без мокроты для активной диагностики туберкулеза (ТБ) имеют высший приоритет для глобальной борьбы с туберкулезом.Мы провели углубленный протеомный анализ с использованием анализа SOMAscan с 4 000 плексами на 1470 образцах сыворотки из семи стран, где ТБ является эндемичным. Все образцы были взяты у пациентов с симптомами и признаками, указывающими на активный туберкулез легких, которые были систематически подтверждены или исключены по результатам посева и клинического наблюдения. Коинфекция ВИЧ присутствовала в 34% проб, а у 25% мазок мокроты был отрицательным. Биомаркеры сывороточного белка были идентифицированы путем отбора стабильности с использованием L1-регуляризованной логистической регрессии и статистики Колмогорова-Смирнова (KS). Наивный байесовский классификатор, использующий шесть маркеров ответа хозяина (модель HR6), включая SYWC, каллистатин, комплемент C9, гельсолин, тестикан-2 и альдолазу C, хорошо показал себя в обучающей выборке (область под кривой чувствительности-специфичности [AUC] 0,94) и в слепом проверочном наборе (AUC 0,92) для различения образцов с туберкулезом и без него. Дифференциальная экспрессия также была высокозначимой ( P <10 -20 ) для ранее описанных маркеров TB, таких как IP-10, LBP, FCG3B и TSP4, а также для многих новых белков, ранее не связанных с TB.Белки с наибольшими изменениями средней кратности были SAA (сывороточный амилоидный белок A), NPS-PLA2 (секретируемая фосфолипаза A2) и CA6 (карбоангидраза 6). Профили целевых продуктов (TPP) для теста на биомаркеры без мокроты для диагностики активного туберкулеза для начала лечения (TPP # 1) и для сортировки по месту жительства или теста на направление (TPP # 2) были опубликованы ВОЗ. Модель HR6 с чувствительностью 90% и специфичностью 80% не дотянула до TPP # 1, но достигла критерия эффективности TPP # 2.
Наивный байесовский классификатор, использующий шесть маркеров ответа хозяина (модель HR6), включая SYWC, каллистатин, комплемент C9, гельсолин, тестикан-2 и альдолазу C, хорошо показал себя в обучающей выборке (область под кривой чувствительности-специфичности [AUC] 0,94) и в слепом проверочном наборе (AUC 0,92) для различения образцов с туберкулезом и без него. Дифференциальная экспрессия также была высокозначимой ( P <10 -20 ) для ранее описанных маркеров TB, таких как IP-10, LBP, FCG3B и TSP4, а также для многих новых белков, ранее не связанных с TB.Белки с наибольшими изменениями средней кратности были SAA (сывороточный амилоидный белок A), NPS-PLA2 (секретируемая фосфолипаза A2) и CA6 (карбоангидраза 6). Профили целевых продуктов (TPP) для теста на биомаркеры без мокроты для диагностики активного туберкулеза для начала лечения (TPP # 1) и для сортировки по месту жительства или теста на направление (TPP # 2) были опубликованы ВОЗ. Модель HR6 с чувствительностью 90% и специфичностью 80% не дотянула до TPP # 1, но достигла критерия эффективности TPP # 2. В заключение мы определили и утвердили сигнатуру с шестью маркерами для активного туберкулеза, которая требует диагностической разработки на платформе рядом с пациентом.
В заключение мы определили и утвердили сигнатуру с шестью маркерами для активного туберкулеза, которая требует диагностической разработки на платформе рядом с пациентом.
ВВЕДЕНИЕ
Туберкулез (ТБ) остается серьезной глобальной проблемой здравоохранения, вызывая самый высокий уровень смертности среди всех инфекционных заболеваний в мире в 2015 году и входит в десятку основных причин всех смертей в мире. Последние отчеты свидетельствуют о том, что, несмотря на значительный прогресс в снижении заболеваемости туберкулезом, по-прежнему есть двое из пяти человек, которым не поставлен диагноз и которые способствуют распространению болезни. Глобальная стратегия ООН по борьбе с туберкулезом нацелена на снижение смертности от туберкулеза на 90% и снижение заболеваемости туберкулезом на 80% к 2030 году (1).Для достижения этих целей критически важны и жизненно важны новые диагностические инструменты для борьбы с эпидемией ТБ (2, 3). Более качественные тесты с использованием образцов, отличных от мокроты, таких как кровь, расширят доступ к диагностическим средствам, которые могут предоставить информацию для лечения и остановить распространение болезни в сообществах (4). Ни один современный диагностический тест не является точным и достаточно дешевым, а существующие тесты часто неадекватны для детей и детей с внелегочными заболеваниями (2–4). Эмпирическое лечение по-прежнему является обычным явлением, особенно когда подозрение на ТБ или риск смерти от нелеченого ТБ высок (5).
Ни один современный диагностический тест не является точным и достаточно дешевым, а существующие тесты часто неадекватны для детей и детей с внелегочными заболеваниями (2–4). Эмпирическое лечение по-прежнему является обычным явлением, особенно когда подозрение на ТБ или риск смерти от нелеченого ТБ высок (5).
Трансформирующая технология, такая как GeneXpert MTB / RIF, уже начала улучшать результаты, но только в сочетании с операционными и инфраструктурными улучшениями (6). Несмотря на всемирное распространение тестов GeneXpert, все еще остаются значительные неудовлетворенные потребности в диагностике. В частности, критически необходимы надежные тесты, не основанные на мокроте, которые можно было бы проводить с небольшими затратами на районном уровне и в медицинских пунктах (7, 8). Для выявления тех, кто нуждается в подтверждающем молекулярном или культуральном тестировании, в том числе тестировании на лекарственную чувствительность (ТЛЧ), на более высоком уровне системы здравоохранения можно использовать чувствительный тест для сортировки. Тест с высокой специфичностью (≥98%) на ТБ позволит начать лечение после положительного результата, но должен сохранять достаточно высокую чувствительность (> 65%), чтобы его можно было использовать в странах, где ТБ является высокоэндемичным.
Тест с высокой специфичностью (≥98%) на ТБ позволит начать лечение после положительного результата, но должен сохранять достаточно высокую чувствительность (> 65%), чтобы его можно было использовать в странах, где ТБ является высокоэндемичным.
Множественные «омические» исследования обнаружили биомаркеры активного ТБ (9–11), включая транскриптомные подходы с использованием секвенирования РНК (12, 13) и обнаружения микроРНК (14), протеомные исследования с помощью масс-спектрометрии для обнаружения пептидов после протеолитического расщепления белков. (15, 16), иммунные исследования, обычно использующие ответы Т-клеток (17), и метаболомику с использованием масс-спектроскопии (18–20).Масс-спектроскопия ограничена соображениями стоимости и чувствительности, может быть трудоемкой и имеет относительно низкую пропускную способность для обнаружения биомаркеров (21). Масс-спектроскопия также технически более сложна в качестве приложения рядом с пациентом по сравнению с простыми форматами анализа, такими как анализ латерального потока или другие сэндвич-тесты с использованием антител или аптамеров для количественного определения конкретных маркеров. Опубликованные на сегодняшний день исследования биомаркеров часто трудно сравнивать, поскольку они сосредоточены на одной конкретной географической области, различаются по количеству субъектов и часто включают контрольные группы населения без представленной информации о латентном статусе ТБ или специфики других инфекционных или воспалительных заболеваний.С 2003 года Фонд инновационных новых диагностических средств (FIND; Женева, Швейцария) курирует высококачественный всемирный банк образцов, чтобы побудить ученых и промышленность к созданию более качественных диагностических тестов. Большое количество образцов сыворотки было любезно предоставлено для нашей работы с помощью анализа SOMAscan. Эта протеомная платформа измеряет> 4000 белков одновременно в небольшом объеме (50 мкл) плазмы или сыворотки, имеет динамический диапазон ∼8 log, средний нижний предел обнаружения 40 фМ и высокую точность (коэффициент вариации <5%). ).Беспристрастные подходы к открытию биомаркеров с помощью SOMAscan привели к диагностическим сигнатурам белков крови для различных заболеваний, поражающих легкие, включая туберкулез (22–24), немелкоклеточный рак легкого (25) и мезотелиому (26).
Опубликованные на сегодняшний день исследования биомаркеров часто трудно сравнивать, поскольку они сосредоточены на одной конкретной географической области, различаются по количеству субъектов и часто включают контрольные группы населения без представленной информации о латентном статусе ТБ или специфики других инфекционных или воспалительных заболеваний.С 2003 года Фонд инновационных новых диагностических средств (FIND; Женева, Швейцария) курирует высококачественный всемирный банк образцов, чтобы побудить ученых и промышленность к созданию более качественных диагностических тестов. Большое количество образцов сыворотки было любезно предоставлено для нашей работы с помощью анализа SOMAscan. Эта протеомная платформа измеряет> 4000 белков одновременно в небольшом объеме (50 мкл) плазмы или сыворотки, имеет динамический диапазон ∼8 log, средний нижний предел обнаружения 40 фМ и высокую точность (коэффициент вариации <5%). ).Беспристрастные подходы к открытию биомаркеров с помощью SOMAscan привели к диагностическим сигнатурам белков крови для различных заболеваний, поражающих легкие, включая туберкулез (22–24), немелкоклеточный рак легкого (25) и мезотелиому (26).
Используя SOMAscan, мы приступили к многоэтапному исследованию, чтобы определить, может ли содержание белка в организме человека быть достаточно устойчивым для решения задач диагностики туберкулеза. Очень сложно обнаружить маркеры патогенного происхождения непосредственно в крови со значимой чувствительностью (27) без использования сложных методологий, таких как масс-спектрометрия, для улучшения обнаружения пептидов, специфичных для Mycobacterium tuberculosis, в переваренных образцах сыворотки (28).
Проведенные нами исследования были разработаны для оценки способности биомаркеров сывороточного белка отличать ТБ-положительных субъектов (подтвержденных положительным посевом мокроты на M. tuberculosis) от не-ТБ субъектов, у которых наблюдаются ТБ-подобные симптомы при наличии и отсутствии ВИЧ. . Представленные здесь результаты демонстрируют, что надежные сигналы от биомаркеров белков-хозяев способны воспроизводимо отличить ТБ от субъектов, не страдающих ТБ. Кроме того, были идентифицированы наборы маркеров, которые коррелировали с ранним ответом на лечение и могли быть полезны для выявления и мониторинга лекарственно-устойчивого туберкулеза (29–31).
РЕЗУЛЬТАТЫ
Субъекты исследования и открытие биомаркеров туберкулеза. С целью улучшения нашего предыдущего исследования биомаркеров фазы I (см. Рис. S1 в дополнительном материале), в ходе которого была получена модель из 9 белков с чувствительностью 80% и специфичностью 84% (Рис. S2) мы использовали более крупную версию SOMAscan и географически более разнообразный набор выборок для обнаружения. Образцы сыворотки крови были собраны у пациентов в нескольких клиниках в семи странах: ЮАР, Перу, Зимбабве, Уганде, Вьетнаме, Колумбии и Бангладеш (Таблица 1).Для обнаружения биомаркеров с помощью SOMAscan было протестировано в общей сложности 252 образца без ТБ и 252 образца с использованием версии, в которой было измерено 4 156 аналитов. Небольшая часть образцов (7,5%) была удалена, поскольку они были гемолизированы ( n = 7), не прошли анализ показателей контроля качества ( n = 15) или были дубликатами ( n = 16). Остальные 466 образцов были признаны пригодными для анализа данных и включали 159 (отрицательный на ТБ, отрицательный результат на ВИЧ), 151 (положительный результат на ТБ, отрицательный результат на ВИЧ, включая 36 отрицательных мазков), 79 (отрицательный на ТБ, положительный результат на ВИЧ) и 77 (положительный результат на ТБ, ВИЧ-положительные, в том числе 23 отрицательных по мазку) образцов. Имелись врожденные демографические различия: пациенты с туберкулезом были моложе ( P = 0,0050), имели более низкий индекс массы тела (ИМТ) ( P = 0,0012) и имели более высокую долю мужчин ( P = 0,0010), чем группа, не болеющая туберкулезом. По замыслу, классы выборки были хорошо сбалансированы с точки зрения ВИЧ-статуса (рис. S3).
Имелись врожденные демографические различия: пациенты с туберкулезом были моложе ( P = 0,0050), имели более низкий индекс массы тела (ИМТ) ( P = 0,0012) и имели более высокую долю мужчин ( P = 0,0010), чем группа, не болеющая туберкулезом. По замыслу, классы выборки были хорошо сбалансированы с точки зрения ВИЧ-статуса (рис. S3).
Демографические и клинические данные участников исследований сывороточных биомаркеров ТБ с использованием анализа SOMAscan 4000 plex (фаза II)
При 5% -ном уровне значимости, скорректированном по Бонферрони, тест Колмогорова-Смирнова (KS) выявил 722/4 156 (17%) белки по-разному экспрессируются в группах ТБ и не-ТБ.Из них 312/722 (43%) были экспрессированы на более высоких уровнях у пациентов с ТБ, чем у пациентов без ТБ. Основные существенно дифференцирующиеся белки изображены на графике вулкана (рис. 1). В дополнение к этому одномерному анализу обнаружения биомаркеров мы применили отбор стабильности с использованием L 1 -регуляризованной модели логистической регрессии. Этот метод позволил включить клинические и демографические метаданные вместе со всеми измерениями белка при сравнении образцов с туберкулезом и не туберкулезом и, как правило, дает наиболее надежные и некоррелированные маркеры (рис.S4).
Этот метод позволил включить клинические и демографические метаданные вместе со всеми измерениями белка при сравнении образцов с туберкулезом и не туберкулезом и, как правило, дает наиболее надежные и некоррелированные маркеры (рис.S4).
дифференциальной экспрессии белков, измеренной в образцах сыворотки от ТБ ( n = 228), по сравнению с субъектами без ТБ ( n = 238). Статистически наиболее значимые маркеры показаны вверху, а белки с наибольшим кратным изменением медианных сигналов — вправо и влево, соответственно. Каллистатин и C9 показаны дважды, потому что в SOMAscan использовались два отдельных SOMAmer для каждого из этих белков, как указано звездочкой.
Верхние маркеры, позволяющие отличить образцы ТБ от образцов, не связанных с ТБ, сведены в Таблицу 2. Каллистатин, SYWC и C9 были тремя основными маркерами, идентифицированными в обоих анализах. Мы подтвердили маркеры, ранее обнаруженные в фазе I, включая маркеры с большими расстояниями KS (каллистатин, C9, NPS-PLA2, IP10, TSP4 и гельсолин) и маркеры с большими изменениями средней кратности (SAA, CRP и карбоангидраза 6). Многие новые маркеры были обнаружены среди большего протеомного содержимого версии SOMAscan с 4000 сплетений по сравнению со сканированием 1,129 plex, используемым в фазе I, включая SYWC, MED-1, FCN1, B4GT6, PGRP-L, антиген CD36, PTGD2, sPLA. (2) -XIII, биотинидаза, люмикан, α1-антитрипсин, FGL1, BMP-1, CD109, кателицидин и альдолаза C.
Многие новые маркеры были обнаружены среди большего протеомного содержимого версии SOMAscan с 4000 сплетений по сравнению со сканированием 1,129 plex, используемым в фазе I, включая SYWC, MED-1, FCN1, B4GT6, PGRP-L, антиген CD36, PTGD2, sPLA. (2) -XIII, биотинидаза, люмикан, α1-антитрипсин, FGL1, BMP-1, CD109, кателицидин и альдолаза C.
Белковые маркеры сыворотки, которые отличают ТБ от не-ТБ среди первоначальных подозреваемых на ТБ, ранжируются по статистике KS a
Новым главным маркером, более распространенным при ТБ, чем в образцах без ТБ, был SYWC, гамма-интерферон ( IFN-γ) -индуцируемая Trp-тРНК-синтетаза, связанная со стрессовой реакцией. Идентичность белков HR6 была подтверждена методом аффинного захвата (вытягивания) из сыворотки с использованием иммобилизованных на гранулах модифицированных аптамеров с медленной скоростью (SOMAmers), как описано в другом исследовании (27).Ограниченная чувствительность этого метода позволяла отбирать сывороточные белки только от среднего до высокого, включая каллистатин, C9, гельсолин и SYWC (рис. S5).
S5).
Моделирование и проверка сигнатуры маркера хозяина для активного туберкулеза у взрослых. Идентификация дополнительных сильных сывороточных белковых маркеров туберкулеза побудила попытаться построить улучшенную модель с повышенной чувствительностью и специфичностью и, возможно, меньшим количеством белков по сравнению с моделью HR9 (рис. S2). Выборки были случайным образом разделены на обучающий набор, содержащий 80% образцов ( n = 190 без ТБ по сравнению с n = 181 ТБ) и тестовый набор, содержащий оставшиеся 20% образцов ( n = 48 без -ТБ против n = 47 ТБ), которые использовались для независимой оценки производительности предварительной модели.Для построения и установления моделей использовалась только обучающая выборка. Кандидаты в маркеры ТБ, которые были связаны с демографическими переменными или клиническими параметрами, отличными от статуса ТБ, были отклонены. Примерами были CCL28, который был хорошим маркером туберкулеза в образцах из Южной Африки, но не из других мест, и sCD163, который отличал туберкулез от туберкулеза только в ВИЧ-положительной субпопуляции (рис. S6). Кроме того, мы требовали, чтобы кандидаты в маркеры имели хорошую корреляцию измерений между сывороткой и плазмой, чтобы последующие модели можно было применять к любому типу матрицы образца.Наконец, кандидаты на биомаркеры ТБ лишались приоритета, если они демонстрировали чрезвычайно узкое распределение сигналов, что делало их менее устойчивыми по сравнению со стандартной гибридизацией и процедурами медианной нормализации, обычно применяемыми к данным SOMAscan. С использованием этих критериев были построены базовые модели с последующим либо обратным исключением, либо прямым отбором с использованием подмножества маркеров, показанных в таблице 2. Сравнение результатов перекрестной проверки характеристик модели кандидата с целью максимизировать любую площадь под кривой зависимости концентрации от времени ( AUC), специфичность или чувствительность и специфичность привели к окончательной наивной байесовской модели с использованием гауссовских плотностей, специфичных для класса, с надежными оценками параметров.
S6). Кроме того, мы требовали, чтобы кандидаты в маркеры имели хорошую корреляцию измерений между сывороткой и плазмой, чтобы последующие модели можно было применять к любому типу матрицы образца.Наконец, кандидаты на биомаркеры ТБ лишались приоритета, если они демонстрировали чрезвычайно узкое распределение сигналов, что делало их менее устойчивыми по сравнению со стандартной гибридизацией и процедурами медианной нормализации, обычно применяемыми к данным SOMAscan. С использованием этих критериев были построены базовые модели с последующим либо обратным исключением, либо прямым отбором с использованием подмножества маркеров, показанных в таблице 2. Сравнение результатов перекрестной проверки характеристик модели кандидата с целью максимизировать любую площадь под кривой зависимости концентрации от времени ( AUC), специфичность или чувствительность и специфичность привели к окончательной наивной байесовской модели с использованием гауссовских плотностей, специфичных для класса, с надежными оценками параметров. Эта оптимальная модель содержала 6 белков ответа хозяина (модель HR6): SYWC, каллистатин, C9, гельсолин, тестикан-2 и альдолаза C. Четыре из шести биомаркеров были частью предыдущей модели HR9 из фазы I, а SYWC и альдолаза. C были обнаружены в более крупном SOMAscan с 4000 плексами, используемом в фазе II, и обеспечили дополнительную чувствительность и специфичность модели. В обучающем наборе модель HR6 показала AUC 0,93 (95% доверительный интервал [95% ДИ], 0,90, 0,95), что было немного лучше, чем производительность предыдущей модели HR9, адаптированной к новым данным (AUC 0 .89 [95% ДИ 0,85, 0,92]). Характеристики модели были подтверждены в наборе для испытаний на удержание 20% из 95 образцов, которые не использовались для построения модели (AUC 0,88 [95% ДИ, 0,79, 0,94]). Важно отметить, что производительность модели существенно не различалась при стратификации по стране, полу или возрасту (данные не показаны). Модель HR6 также применялась к набору проверок специфичности, который включал 82 образца (40 из Канады и 42 из Испании) от пациентов, не страдающих туберкулезом, с легочными заболеваниями, такими как хроническая обструктивная болезнь легких, бронхоэктазы, астма, пневмония, бронхит, одышка, тальк.
Эта оптимальная модель содержала 6 белков ответа хозяина (модель HR6): SYWC, каллистатин, C9, гельсолин, тестикан-2 и альдолаза C. Четыре из шести биомаркеров были частью предыдущей модели HR9 из фазы I, а SYWC и альдолаза. C были обнаружены в более крупном SOMAscan с 4000 плексами, используемом в фазе II, и обеспечили дополнительную чувствительность и специфичность модели. В обучающем наборе модель HR6 показала AUC 0,93 (95% доверительный интервал [95% ДИ], 0,90, 0,95), что было немного лучше, чем производительность предыдущей модели HR9, адаптированной к новым данным (AUC 0 .89 [95% ДИ 0,85, 0,92]). Характеристики модели были подтверждены в наборе для испытаний на удержание 20% из 95 образцов, которые не использовались для построения модели (AUC 0,88 [95% ДИ, 0,79, 0,94]). Важно отметить, что производительность модели существенно не различалась при стратификации по стране, полу или возрасту (данные не показаны). Модель HR6 также применялась к набору проверок специфичности, который включал 82 образца (40 из Канады и 42 из Испании) от пациентов, не страдающих туберкулезом, с легочными заболеваниями, такими как хроническая обструктивная болезнь легких, бронхоэктазы, астма, пневмония, бронхит, одышка, тальк. -индуцированное заболевание легких или эмфизема легких.Эти образцы были от более старшего населения (средний возраст 64,5 года; диапазон от 19 до 93 лет) и включали 22 пациента с туберкулезом в анамнезе и 9 пациентов с подтвержденной нетуберкулезной микобактериальной (НТМ) инфекцией. HR6 показал себя достаточно хорошо в этом наборе заражений (специфичность 93%), с 6 ложными срабатываниями, включая одну грубую ошибочную классификацию (логарифм шансы> 5) и только один из девяти пациентов с НТМ, ошибочно классифицированных как ТБ (логарифм шансы 1,67), что указывает на то, что на специфичность модели HR6 не влияло присутствие NTM.Аналогичным образом модель HR6 хорошо себя показала в контрольном наборе из 168 образцов из здоровой нормальной популяции, для которой у нас были данные SOMAscan в файле (специфичность> 98%), с 3 слабыми ложноположительными результатами. Разделение образцов TB и non-TB показано графиками кумулятивной функции распределения (CDF) для каждого из маркеров HR6, показанных на рис.
-индуцированное заболевание легких или эмфизема легких.Эти образцы были от более старшего населения (средний возраст 64,5 года; диапазон от 19 до 93 лет) и включали 22 пациента с туберкулезом в анамнезе и 9 пациентов с подтвержденной нетуберкулезной микобактериальной (НТМ) инфекцией. HR6 показал себя достаточно хорошо в этом наборе заражений (специфичность 93%), с 6 ложными срабатываниями, включая одну грубую ошибочную классификацию (логарифм шансы> 5) и только один из девяти пациентов с НТМ, ошибочно классифицированных как ТБ (логарифм шансы 1,67), что указывает на то, что на специфичность модели HR6 не влияло присутствие NTM.Аналогичным образом модель HR6 хорошо себя показала в контрольном наборе из 168 образцов из здоровой нормальной популяции, для которой у нас были данные SOMAscan в файле (специфичность> 98%), с 3 слабыми ложноположительными результатами. Разделение образцов TB и non-TB показано графиками кумулятивной функции распределения (CDF) для каждого из маркеров HR6, показанных на рис. 2.
2.
Графики кумулятивной функции распределения (CDF) шести маркеров хозяина, составляющих HR6 модель (SYWC, каллистатин, комплемент C9, гельсолин, альдолаза C и тестикан-2) в обучающей выборке из 181 ТБ (красный) по сравнению с 190 образцами без ТБ (синий).Также показаны измерения в образцах из Испании и Канады (зеленый цвет), полученных от пациентов с заболеваниями легких, отличными от туберкулеза.
Модель HR6 была заблокирована и предоставлена FIND перед тестированием слепого проверочного набора из 216 образцов, снова включая несколько реплицируемых образцов ( n = 19), чтобы обеспечить возможность объединения отдельных наборов данных. Прогнозы с использованием модели HR6 были сделаны для 212 образцов, за исключением трех гемолизированных образцов и одного неудачного анализа, а рассчитанные логарифмические шансы на ТБ были представлены в FIND.Раскрытие метаданных выявило восемь образцов от субъектов с неполными клиническими данными, которые были исключены из анализа. В 204 оставшихся проверочных образцах производительность модели HR6 показала AUC 0,87 (95% ДИ, 0,81, 0,91), что находится в пределах ожидаемой чувствительности и специфичности в рабочих точках Байеса, основанных на оценках начальной загрузки соответствующей эмпирической достоверности 95%. интервалы. Кривые рабочих характеристик приемника (ROC) для 670 выборок, стратифицированных обучающими, тестовыми и слепыми проверочными наборами, показаны на рис.3А.
В 204 оставшихся проверочных образцах производительность модели HR6 показала AUC 0,87 (95% ДИ, 0,81, 0,91), что находится в пределах ожидаемой чувствительности и специфичности в рабочих точках Байеса, основанных на оценках начальной загрузки соответствующей эмпирической достоверности 95%. интервалы. Кривые рабочих характеристик приемника (ROC) для 670 выборок, стратифицированных обучающими, тестовыми и слепыми проверочными наборами, показаны на рис.3А.
Кривые ROC для первоначальной оценки производительности модели HR6, чтобы отличить ТБ от образцов сыворотки, не связанных с ТБ. (A) Все образцы фазы II ( n = 670), стратифицированные путем обучения, тестирования и слепой проверки. (B) Стратификация по выборочной оценке достоверности метаданных. Подавляющее большинство выборок ( n = 620; 92,5%) были связаны с надежными метаданными, которые обеспечивали высокий уровень уверенности в их истинной классификации, в то время как небольшая подгруппа выборок ( n = 50; 7. 5%) имели менее убедительные метаданные. (C) Производительность модели HR6 в выборках фазы II с метаданными высокой достоверности, стратифицированными по обучению, тестированию и слепому набору проверки. Оптимальная рабочая точка для кривой HR6 ROC была при чувствительности 90% и специфичности 80%, как указано черной звездочкой. Для справки: минимальные требования к производительности для TPP № 1 (тест для диагностики активного ТБ) и TPP № 2 (для сортировки / направления) показаны синими и зелеными звездочками соответственно.
5%) имели менее убедительные метаданные. (C) Производительность модели HR6 в выборках фазы II с метаданными высокой достоверности, стратифицированными по обучению, тестированию и слепому набору проверки. Оптимальная рабочая точка для кривой HR6 ROC была при чувствительности 90% и специфичности 80%, как указано черной звездочкой. Для справки: минимальные требования к производительности для TPP № 1 (тест для диагностики активного ТБ) и TPP № 2 (для сортировки / направления) показаны синими и зелеными звездочками соответственно.
Ошибочные классификации, несоответствия в данных и оценки качества выборки.Модель HR6 произвела несколько грубых ошибок в классификации наборов для обучения, тестирования и проверки, что потребовало дальнейшего анализа протеомных данных и связанных с ними метаданных. Уровень канцелярских ошибок при сборе данных, маркировке, аликвоте и управлении базами данных оказался низким для 670 образцов, проанализированных на этапе II. Сигналы для белков, специфичных для пола (ФСГ, ЛГ и ПСА), в целом очень соответствовали метаданным. Один субъект имел несоответствующие записи по возрасту и полу по сравнению с метаданными для отдельной аликвоты сыворотки от того же субъекта, предоставленной с разницей в 2 года.Другой субъект, несомненно, был беременным на основании протеомного профиля, но был отмечен как мужчина. Среди в общей сложности 64 субъектов, для которых дублированные аликвоты были получены и протестированы отдельно во время этапов I и II анализов, 58 показали высоко согласованные протеомные измерения (rho> 0,98), типичные для повторений от одного и того же субъекта. Однако другие 6 парных выборок явно противоречили друг другу (rho <0,90), что обычно наблюдается у неродственных людей, и четыре из них были образцами, которые были перемаркированы в целях ослепления для использования в проверочной выборке.Мы также повторно проверили качество и количество метаданных, предоставленных с образцами, и разработали оценку, чтобы определить, насколько мы уверены в истинном диагнозе ТБ (баллы: 1 - идеально; 0,75 - хорошо; 0,5 - неполные данные; 0,25 - сомнительно; 0).
Один субъект имел несоответствующие записи по возрасту и полу по сравнению с метаданными для отдельной аликвоты сыворотки от того же субъекта, предоставленной с разницей в 2 года.Другой субъект, несомненно, был беременным на основании протеомного профиля, но был отмечен как мужчина. Среди в общей сложности 64 субъектов, для которых дублированные аликвоты были получены и протестированы отдельно во время этапов I и II анализов, 58 показали высоко согласованные протеомные измерения (rho> 0,98), типичные для повторений от одного и того же субъекта. Однако другие 6 парных выборок явно противоречили друг другу (rho <0,90), что обычно наблюдается у неродственных людей, и четыре из них были образцами, которые были перемаркированы в целях ослепления для использования в проверочной выборке.Мы также повторно проверили качество и количество метаданных, предоставленных с образцами, и разработали оценку, чтобы определить, насколько мы уверены в истинном диагнозе ТБ (баллы: 1 - идеально; 0,75 - хорошо; 0,5 - неполные данные; 0,25 - сомнительно; 0). , нет данных), как подробно показано на фиг. S7A. По сути, для получения хорошего показателя достоверности метаданных необходимы согласованные данные о посеве как минимум двух мокроты на ТБ и полная информация о последующих наблюдениях для субъектов, не страдающих ТБ. В целом, метаданные выборки были очень хорошими: 95% образцов, не относящихся к туберкулезу, и 89% выборок с туберкулезом имели балл ≥0.75. Выборки с низкими показателями достоверности метаданных чаще встречались среди ошибочных классификаций HR6, и, следовательно, модель HR6 работала намного лучше в выборках с высокой степенью достоверности (AUC 0,93 [95% ДИ, 0,91, 0,95]), чем в выборках с низкой достоверностью. (AUC 0,60 [95% ДИ, 0,42, 0,75]), как показано на кривых ROC (фиг. 3B). Исключение 50 образцов с сомнительными метаданными и удаление 4 противоречивых дубликатов привело к небольшому увеличению производительности модели в оставшихся 616 образцах (рис.3С). В наборе слепой проверки ( n = 177) AUC увеличилась с 0,87 (95% ДИ, 0,81, 0,91) до 0,92 (95% ДИ, 0,87, 0,96) в образцах с высокой степенью достоверности.
, нет данных), как подробно показано на фиг. S7A. По сути, для получения хорошего показателя достоверности метаданных необходимы согласованные данные о посеве как минимум двух мокроты на ТБ и полная информация о последующих наблюдениях для субъектов, не страдающих ТБ. В целом, метаданные выборки были очень хорошими: 95% образцов, не относящихся к туберкулезу, и 89% выборок с туберкулезом имели балл ≥0.75. Выборки с низкими показателями достоверности метаданных чаще встречались среди ошибочных классификаций HR6, и, следовательно, модель HR6 работала намного лучше в выборках с высокой степенью достоверности (AUC 0,93 [95% ДИ, 0,91, 0,95]), чем в выборках с низкой достоверностью. (AUC 0,60 [95% ДИ, 0,42, 0,75]), как показано на кривых ROC (фиг. 3B). Исключение 50 образцов с сомнительными метаданными и удаление 4 противоречивых дубликатов привело к небольшому увеличению производительности модели в оставшихся 616 образцах (рис.3С). В наборе слепой проверки ( n = 177) AUC увеличилась с 0,87 (95% ДИ, 0,81, 0,91) до 0,92 (95% ДИ, 0,87, 0,96) в образцах с высокой степенью достоверности. Перекрестная таблица результатов теста HR6 для 616 образцов с помощью эталонного стандарта FIND показала, что не было статистической разницы в производительности модели HR6 между обучающими и слепыми проверочными наборами (таблица 3). Логарифмические распределения шансов для этих 616 выборок, иллюстрирующие ошибочную классификацию, показаны на рис.S7B. Производительность модели HR6 была независимо рассчитана и подтверждена персоналом FIND.
Перекрестная таблица результатов теста HR6 для 616 образцов с помощью эталонного стандарта FIND показала, что не было статистической разницы в производительности модели HR6 между обучающими и слепыми проверочными наборами (таблица 3). Логарифмические распределения шансов для этих 616 выборок, иллюстрирующие ошибочную классификацию, показаны на рис.S7B. Производительность модели HR6 была независимо рассчитана и подтверждена персоналом FIND.
Сопоставление результатов теста HR6 SOMAscan с помощью эталонного теста для всех обучающих, тестовых и слепых проверочных образцов фазы II с полными метаданными a
Влияние коинфекции ВИЧ и статуса мазка на производительность модели HR6. Повторяющиеся образцы ( n = 47), которые были частью как открытия, так и слепой проверки, были удалены, а оставшийся набор уникальных образцов с высокой степенью достоверности ( n = 569) был использован для более подробного расчета чувствительности и специфика модели HR6 в разных предметных подгруппах. Кривые ROC, позволяющие отличить ТБ ( n = 262) от субъектов, не страдающих ТБ ( n = 307), стратифицированные по ВИЧ-статусу, показали лишь незначительное снижение точности теста для ВИЧ-положительных субъектов ТБ ( n = 85; AUC 0,93 [95% ДИ, 0,87, 0,96]) по сравнению с ВИЧ-отрицательными пациентами с ТБ ( n = 177; AUC 0,94 [95% ДИ, 0,92, 0,96]), но более резкое падение у пациентов с отрицательным мазком мокроты ( n = 52; AUC 0,87 [95% ДИ, 0,79, 0,92]) по сравнению с пациентами с ТБ с положительным мазком ( n = 210; AUC 0.95 [95% ДИ, 0,93, 0,97]) (рис. S7C). Среди 569 уникальных образцов была 21 грубая неправильная классификация, в том числе 10 ложноотрицательных результатов (логарифм <−5), 6 из которых были получены от субъектов с ТБ с отрицательным мазком мокроты. Чувствительность и специфичность HR6 для выявления активного легочного ТБ во всех уникальных образцах ( n = 569) суммированы в таблице 4, стратифицированной по коинфекции ВИЧ и статусу мазка.
Кривые ROC, позволяющие отличить ТБ ( n = 262) от субъектов, не страдающих ТБ ( n = 307), стратифицированные по ВИЧ-статусу, показали лишь незначительное снижение точности теста для ВИЧ-положительных субъектов ТБ ( n = 85; AUC 0,93 [95% ДИ, 0,87, 0,96]) по сравнению с ВИЧ-отрицательными пациентами с ТБ ( n = 177; AUC 0,94 [95% ДИ, 0,92, 0,96]), но более резкое падение у пациентов с отрицательным мазком мокроты ( n = 52; AUC 0,87 [95% ДИ, 0,79, 0,92]) по сравнению с пациентами с ТБ с положительным мазком ( n = 210; AUC 0.95 [95% ДИ, 0,93, 0,97]) (рис. S7C). Среди 569 уникальных образцов была 21 грубая неправильная классификация, в том числе 10 ложноотрицательных результатов (логарифм <−5), 6 из которых были получены от субъектов с ТБ с отрицательным мазком мокроты. Чувствительность и специфичность HR6 для выявления активного легочного ТБ во всех уникальных образцах ( n = 569) суммированы в таблице 4, стратифицированной по коинфекции ВИЧ и статусу мазка. Чтобы быть еще более строгими, мы определили производительность модели после исключения всех обучающих выборок, тем самым ограничив анализ только комбинированной тестовой и проверочной выборками ( n = 216).Кривые ROC, чтобы отличить ТБ ( n = 96) от субъектов, не страдающих ТБ ( n = 120), снова были стратифицированы по ВИЧ-инфекции и статусу мазка (рис. S7D). Показатели HR6 снизились у ВИЧ-положительных субъектов с ТБ ( n = 30; AUC 0,88 [95% ДИ, 0,75, 0,95]) по сравнению с ВИЧ-отрицательными пациентами с ТБ ( n = 66; AUC 0,94 [95% ДИ 0,89, 0,97]) и при ТБ с отрицательным мазком ( n = 11; AUC 0,77 [95% ДИ, 0,53, 0,91]) по сравнению с пациентами с ТБ с положительным мазком ( n = 85; AUC 0.94 [95% ДИ 0,89, 0,97]).
Чтобы быть еще более строгими, мы определили производительность модели после исключения всех обучающих выборок, тем самым ограничив анализ только комбинированной тестовой и проверочной выборками ( n = 216).Кривые ROC, чтобы отличить ТБ ( n = 96) от субъектов, не страдающих ТБ ( n = 120), снова были стратифицированы по ВИЧ-инфекции и статусу мазка (рис. S7D). Показатели HR6 снизились у ВИЧ-положительных субъектов с ТБ ( n = 30; AUC 0,88 [95% ДИ, 0,75, 0,95]) по сравнению с ВИЧ-отрицательными пациентами с ТБ ( n = 66; AUC 0,94 [95% ДИ 0,89, 0,97]) и при ТБ с отрицательным мазком ( n = 11; AUC 0,77 [95% ДИ, 0,53, 0,91]) по сравнению с пациентами с ТБ с положительным мазком ( n = 85; AUC 0.94 [95% ДИ 0,89, 0,97]).
Чувствительность и специфичность HR6 для активного выявления легочного ТБ, стратифицированная по коинфекции ВИЧ и статусу мазка
Разработка целевого панельного анализа ТБ и альтернативных моделей. Целевая панель содержала небольшое подмножество всего меню SOMAscan, включая Зонды SOMAmer для маркеров TB, показанные в таблице 2. Это позволило считывать показания на гораздо меньшем массиве слайдов с низкой плотностью (Applied Microarrays, Inc.), что привело к более экономичному формату анализа.Панельный анализ проводился как в полуавтоматическом, так и в ручном формате с репрезентативной подгруппой ТБ ( n = 39) и не-ТБ ( n = 35) образцов. Биомаркеры туберкулеза, обнаруженные при полном сканировании SOMA, были подтверждены на специальной панели с учетом как статистики KS, так и кратного изменения медианы между образцами TB и non-TB (рис. 4).
Целевая панель содержала небольшое подмножество всего меню SOMAscan, включая Зонды SOMAmer для маркеров TB, показанные в таблице 2. Это позволило считывать показания на гораздо меньшем массиве слайдов с низкой плотностью (Applied Microarrays, Inc.), что привело к более экономичному формату анализа.Панельный анализ проводился как в полуавтоматическом, так и в ручном формате с репрезентативной подгруппой ТБ ( n = 39) и не-ТБ ( n = 35) образцов. Биомаркеры туберкулеза, обнаруженные при полном сканировании SOMA, были подтверждены на специальной панели с учетом как статистики KS, так и кратного изменения медианы между образцами TB и non-TB (рис. 4).
Согласование сфокусированной ТВ-панели с полным сканированием SOMA на 4000 сплетений. (A) Расстояния KS для 40 релевантных маркеров на основе полного анализа SOMAscan обучающих выборок по сравнению с целевым панельным анализом TB для подмножества TB ( n = 39) и не-TB ( n = 35) образцов.Три верхних маркера на основе статистики KS (каллистатин, SYWC и C9) показаны светлыми кружками. (B) Среднее кратное изменение для тех же 40 маркеров, основанное на измерениях на полных данных SOMAscan всех уникальных образцов фазы II и в панельном анализе подмножеств TB ( n = 39) и не-TB ( n = 35) образцов соответственно. Маркеры с большими изменениями средней кратности (SAA, NPS-PLA2, IP-10 и CA6) показаны светлыми кружками.
(B) Среднее кратное изменение для тех же 40 маркеров, основанное на измерениях на полных данных SOMAscan всех уникальных образцов фазы II и в панельном анализе подмножеств TB ( n = 39) и не-TB ( n = 35) образцов соответственно. Маркеры с большими изменениями средней кратности (SAA, NPS-PLA2, IP-10 и CA6) показаны светлыми кружками.
HR6 содержал оптимальное количество устойчивых маркеров для достижения желаемой производительности в обучающем наборе (AUC 0.93). Для сравнения: гораздо более простые модели только с двумя статистически наиболее значимыми маркерами (каллистатин и SYWC) или с четырьмя белками, показывающими наибольшие кратные изменения между образцами ТБ и нетуберкулезом (SAA, NPS-PLA2, IP-10 и CA6) имели явно худшие показатели в той же группе (рис. S8).
ОБСУЖДЕНИЕ
Мы провели большое многофазное исследование для выявления и количественной оценки сывороточных белковых маркеров, указывающих на активный туберкулез легких. Анализ SOMAscan предоставил чувствительную высокопроизводительную платформу для быстрого и точного проведения протеомных измерений более 4000 белков в почти 1500 образцах сыворотки. Следуя стандартным подходам к биоинформатике, были идентифицированы наиболее значимые и надежные биомаркеры, которые впоследствии были использованы для построения небольшой протеомной модели (HR6), которая затем была успешно проверена на слепых проверочных образцах.
Следуя стандартным подходам к биоинформатике, были идентифицированы наиболее значимые и надежные биомаркеры, которые впоследствии были использованы для построения небольшой протеомной модели (HR6), которая затем была успешно проверена на слепых проверочных образцах.
Дополнительные силы к нашим результатам заключается в том, что мы обнаружили много ранее открытых белков (23), и протеомные данные биологически и клинически правдоподобны. Биологические категории маркеров включают иммунитет и защиту хозяина (C9, FCG3B, кателицидин, LBP и FCRL1), ремоделирование сосудов (каллистатин и TSP4), внеклеточный матрикс и ремоделирование ткани (гельсолин, COMP, люмикан, тестикан-2 и CD36. антиген), транспорт и регуляция липидов (фосфолипазы), коагуляция и комплемент (C1QT1), протеазы (MMP-1, MMP-2, CNDP1 и BMP-1) и распознавание бактериями (PGRP-L).Среди главных маркеров — каллистатин, и мы ранее сообщали, что уровни каллистатина увеличились после 8-недельной интенсивной фазы противотуберкулезной терапии (23), что согласуется с нашими данными о более низких уровнях каллистатина при ТБ по сравнению с сывороткой, не связанной с ТБ. Каллистатин, ингибитор протеазы калликреина, обладает антиангиогенными, антиоксидантными, антиапоптотическими и противовоспалительными свойствами, а его концентрация в крови снижается при бактериальной пневмонии (32). SYWC представляет собой интересный белок, который ранее не описывался как предполагаемый диагностический маркер, и было обнаружено, что он обладает высокой степенью различения в сыворотке крови активного ТБ человека.SYWC — это индуцируемая интерфероном гамма цитоплазматическая форма триптофанил-тРНК синтетазы (также называемая WARS или TrpRS). Триптофан необходим для M. tuberculosis (33), особенно для синтеза белка оболочки, важного для выживания в макрофагах (34). Повышенная регуляция SYWC в организме хозяина приводит к ограничению свободного триптофана, а истощение триптофана является летальным для M. tuberculosis (35). Совсем недавно было обнаружено, что SYWC активируется в клетках THP-1 человека, инфицированных M. tuberculosis (36).В качестве отдельного механизма во время инфекции ТБ триптофан часто становится ограничивающим из-за его превращения в кинуренин индоламин-2,3-диоксигеназой (37).
Каллистатин, ингибитор протеазы калликреина, обладает антиангиогенными, антиоксидантными, антиапоптотическими и противовоспалительными свойствами, а его концентрация в крови снижается при бактериальной пневмонии (32). SYWC представляет собой интересный белок, который ранее не описывался как предполагаемый диагностический маркер, и было обнаружено, что он обладает высокой степенью различения в сыворотке крови активного ТБ человека.SYWC — это индуцируемая интерфероном гамма цитоплазматическая форма триптофанил-тРНК синтетазы (также называемая WARS или TrpRS). Триптофан необходим для M. tuberculosis (33), особенно для синтеза белка оболочки, важного для выживания в макрофагах (34). Повышенная регуляция SYWC в организме хозяина приводит к ограничению свободного триптофана, а истощение триптофана является летальным для M. tuberculosis (35). Совсем недавно было обнаружено, что SYWC активируется в клетках THP-1 человека, инфицированных M. tuberculosis (36).В качестве отдельного механизма во время инфекции ТБ триптофан часто становится ограничивающим из-за его превращения в кинуренин индоламин-2,3-диоксигеназой (37). Этот фермент фактически входил в число первых 5% маркеров в нашем анализе SOMAscan и действительно был активирован при ТБ (KS 0,33), хотя и не так сильно, как SYWC. Обнаружение нецитокиновых маркеров, таких как SYWC, расширяет биологию протеомики ТБ, которая может быть менее подвержена изменению другими сопутствующими заболеваниями.
Этот фермент фактически входил в число первых 5% маркеров в нашем анализе SOMAscan и действительно был активирован при ТБ (KS 0,33), хотя и не так сильно, как SYWC. Обнаружение нецитокиновых маркеров, таких как SYWC, расширяет биологию протеомики ТБ, которая может быть менее подвержена изменению другими сопутствующими заболеваниями.
Наши данные по биомаркерам в целом хорошо согласуются с результатами исследования Achkar et al.с использованием сыворотки, собранной в больницах Нью-Йорка, где маркеры активного туберкулеза были идентифицированы с помощью жидкостной хроматографии и масс-спектрометрии (15). Наши маркеры и сигнатуры хорошо проявили себя у ВИЧ-инфицированных, несмотря на то, что ВИЧ сильно изменяет легочную среду (38). Примерно одна треть нашей исследуемой популяции была ВИЧ-положительной, и открытие биомаркеров и построение модели были выполнены в полном наборе выборки, хотя мы не приоритизировали маркеры ТБ, на которые повлиял ВИЧ-статус. Напротив, в исследовании Achkar (15) представлены отдельные списки сывороточных маркеров и сигнатур для ВИЧ-отрицательных и ВИЧ-положительных групп.
ТБ эволюционировал вместе с людьми в течение тысяч лет, и наш широкий протеомный подход к обнаружению интересных биологических функций может дать важные сведения о том, как этот патоген стал таким успешным (39). Более подробные белок-белковые взаимодействия в отношении маркеров сыворотки, описанные в этом исследовании, можно увидеть с помощью функции базы данных String, показанной на рис. S9 в дополнительном материале. Интересно, что ранее сообщалось о нескольких маркерах легочного ТБ как части 7-белкового классификатора рака легких, включая C9, CRP и карбоангидразу (25), а C9 и каллистатин были частью 13-маркерного признака мезотелиомы. (26).Это совпадение неудивительно, учитывая сходство в отношении хронической природы этих заболеваний легких, включая общие пути защиты хозяина — иммунные ответы и восстановление и ремоделирование тканей. Однако наши дополнительные маркеры добавляют специфичность для инфекции M. tuberculosis модели HR6.
Следующая цель — разработать быструю, недорогую и практичную платформу, которую можно было бы развернуть в соответствующих условиях. Профили целевых продуктов (TTP) для диагностики туберкулеза для образцов, отличных от мокроты, были определены в качестве приоритетных при участии ключевых экспертов (40) и впоследствии были приняты и опубликованы ВОЗ (41).Первый TPP предназначен для быстрого диагностического теста на основе биомаркеров с очень высокой специфичностью (98%), который может использоваться как автономный или подтверждающий тест. Второй TPP предназначен для сортировки или перенаправления, который имеет высокую чувствительность (от 90 до 95%), но умеренную специфичность (от 70 до 80%) и может использоваться для выявления людей с предположительным туберкулезом или для исключения туберкулеза в случае отрицательного результата. Как видно из кривых ROC, представленных на рис. 3C, модель HR6 соответствует критериям эффективности для теста направления / сортировки, но не для теста обнаружения ТБ.Оптимальной рабочей точкой модели HR6 была 90% чувствительность и 80% специфичность в целом, хотя точность теста была ниже у ВИЧ-положительных пациентов по сравнению с ВИЧ-отрицательными пациентами ТБ, а также была ниже при ТБ с отрицательным мазком, чем при ТБ с положительным мазком .
Профили целевых продуктов (TTP) для диагностики туберкулеза для образцов, отличных от мокроты, были определены в качестве приоритетных при участии ключевых экспертов (40) и впоследствии были приняты и опубликованы ВОЗ (41).Первый TPP предназначен для быстрого диагностического теста на основе биомаркеров с очень высокой специфичностью (98%), который может использоваться как автономный или подтверждающий тест. Второй TPP предназначен для сортировки или перенаправления, который имеет высокую чувствительность (от 90 до 95%), но умеренную специфичность (от 70 до 80%) и может использоваться для выявления людей с предположительным туберкулезом или для исключения туберкулеза в случае отрицательного результата. Как видно из кривых ROC, представленных на рис. 3C, модель HR6 соответствует критериям эффективности для теста направления / сортировки, но не для теста обнаружения ТБ.Оптимальной рабочей точкой модели HR6 была 90% чувствительность и 80% специфичность в целом, хотя точность теста была ниже у ВИЧ-положительных пациентов по сравнению с ВИЧ-отрицательными пациентами ТБ, а также была ниже при ТБ с отрицательным мазком, чем при ТБ с положительным мазком . Частота ошибок классификации HR6 у субъектов из стран с высоким и средним бременем болезни была намного выше, чем диагностические ошибки у субъектов из Испании и Канады, где бремя туберкулеза ниже, что позволяет предположить, что наши сигнатуры чувствительны к субклиническим зарождающимся заболеваниям.Скрытая инфекция все чаще рассматривается как спектр заболеваний, и интересно предположить, что некоторые из ложноположительных результатов связаны с субклиническим заболеванием (42). Хотя мы не обучали эту модель в районах с низким бременем ТБ, в большой коллекции выборок здоровых субъектов из США не было серьезных ошибок классификации.
Частота ошибок классификации HR6 у субъектов из стран с высоким и средним бременем болезни была намного выше, чем диагностические ошибки у субъектов из Испании и Канады, где бремя туберкулеза ниже, что позволяет предположить, что наши сигнатуры чувствительны к субклиническим зарождающимся заболеваниям.Скрытая инфекция все чаще рассматривается как спектр заболеваний, и интересно предположить, что некоторые из ложноположительных результатов связаны с субклиническим заболеванием (42). Хотя мы не обучали эту модель в районах с низким бременем ТБ, в большой коллекции выборок здоровых субъектов из США не было серьезных ошибок классификации.
Что касается достижения TPP, особенно для теста на выявление туберкулеза со специфичностью ≥98%, точность «стандарта истинности» имеет решающее значение, чтобы избежать ошибочно классифицированных образцов, которые снизят кажущуюся производительность новой модели.Очевидно, что модель HR6 показала гораздо лучшие результаты в подмножестве выборок с высокими показателями достоверности метаданных по сравнению с подмножеством образцов (7,5%) с низкими показателями достоверности.
Мы надеемся, что этот тест может быть разработан и развернут для активного выявления случаев заболевания на уровне системы здравоохранения, который связан с надлежащим ведением случаев и возможностями лечения для максимального воздействия. В этой области была проделана большая работа по анализу воздействия GeneXpert, который открывает путь для новой и точной диагностики туберкулеза (6).Хотя анализ Xpert MTB / RIF — важный шаг вперед, он плохо работает с сывороткой или кровью. Тест LAM в моче хорошо работает только у ВИЧ-положительных субъектов с низким количеством клеток CD4. Мы надеемся, что наш тест будет более чувствительным по спектру CD4. Такой тест мог бы быть полезным инструментом скрининга с использованием образцов не мокроты во всех слоях пациентов. Более того, панельный тест белковых биомаркеров может увеличить выявление случаев заболевания, сократить задержку диагностики и снизить передачу, если он проводится в контексте функциональной системы здравоохранения (4, 43–46). В случае перехода на более дешевую платформу, ориентированную на пациента, ее можно было бы развернуть в периферийных медицинских пунктах, чтобы охватить наибольшую часть пациентов, обращающихся за помощью, но для достижения этих целей потребуется дополнительное внутреннее или донорское финансирование (47).
В случае перехода на более дешевую платформу, ориентированную на пациента, ее можно было бы развернуть в периферийных медицинских пунктах, чтобы охватить наибольшую часть пациентов, обращающихся за помощью, но для достижения этих целей потребуется дополнительное внутреннее или донорское финансирование (47).
МАТЕРИАЛЫ И МЕТОДЫ
Дизайн исследования и сбор образцов. FIND и ее партнерские сайты в Южной Африке, Перу, Зимбабве, Уганде, Вьетнаме, Колумбии и Бангладеш включили взрослых, у которых были признаки и симптомы ТБ (кашель не менее 2 лет). недели, лихорадка, потеря веса и ночная потливость).Были собраны основные демографические данные, такие как возраст, вес и пол, и клинические метаданные, такие как ВИЧ-статус и, в некоторых случаях, количество клеток CD4 и вирусная нагрузка. Рентгенограммы грудной клетки были выполнены и интерпретированы местными радиологами. Образцы мокроты были получены для кислотостойкого окрашивания, твердых и жидких культур и, иногда, для тестирования GeneXpert MTB / RIF. Субъекты с положительным результатом посева на твердой (Левенштейна-Йенсена) и жидкой (пробирка с индикатором роста микобактерий) средах или на обеих считались подтвержденными случаями ТБ.Пациенты, прошедшие химиотерапию ТБ более 5 дней, были исключены. Критерии исключения ТБ были определены FIND и включали отрицательный посев и мазок мазка, а также исчезновение симптомов при отсутствии специфической терапии ТБ при последующем наблюдении через 2–3 месяца.
Субъекты с положительным результатом посева на твердой (Левенштейна-Йенсена) и жидкой (пробирка с индикатором роста микобактерий) средах или на обеих считались подтвержденными случаями ТБ.Пациенты, прошедшие химиотерапию ТБ более 5 дней, были исключены. Критерии исключения ТБ были определены FIND и включали отрицательный посев и мазок мазка, а также исчезновение симптомов при отсутствии специфической терапии ТБ при последующем наблюдении через 2–3 месяца.
Все образцы сыворотки были получены на исходном уровне, получили уникальный штрих-код и заморожены на месте в аликвотах объемом 0,5 мл перед отправкой в центральное хранилище. Этическое одобрение было получено FIND на местах. Некоторые из образцов были из исходного банка образцов ВОЗ / TDR, который в настоящее время находится в ведении FIND, включая сыворотки из Колумбии и Уганды, а также набор нетуберкулезных образцов из Испании и Канады.Отдельный набор образцов здоровой контрольной сыворотки от Covance, Inc. был доступен в биорепозитории SomaLogic и также был проанализирован. SomaLogic получил только деидентифицированные образцы.
SomaLogic получил только деидентифицированные образцы.
Аликвоты замороженной сыворотки были получены в SomaLogic несколькими партиями для двух фаз исследования (таблица 1; см. Также рис. S1A в дополнительном материале). Фаза I и фаза II проводились с интервалом примерно в 2 года и различались разнообразием географического происхождения образцов и количеством белков, для которых определяли сывороточные уровни.Каждая фаза исследования была разработана таким образом, чтобы иметь начальную стадию открытия биомаркеров и построения модели, которая включала образцы от субъектов, выбранных FIND. Стадия слепой проверки включала отдельную отправку набора образцов, выбранного FIND, без диагностической классификации или клинических метаданных. Ключ для описания класса образцов надежно хранился в штаб-квартире FIND и не передавался SomaLogic до тех пор, пока не были завершены протеомные анализы и диагностические прогнозы для слепых образцов не были депонированы в FIND.
Протеомный анализ (SOMAscan). Была применена версия SOMAscan, использовавшаяся на момент поступления образцов: 1,129-сплетение в фазе I и 4,000-сплетение в фазе II. SOMAscan — это запатентованный мультиплексный рабочий процесс для обнаружения относительного количества сигналов, представляющих белки, распознаваемые реагентами с медленным изменением аптамера (SOMAmer), который более подробно описан в другом месте (22). Вкратце, реагенты SOMAmer для более чем 4000 белков были созданы с помощью процедуры, известной как систематическая эволюция лигандов путем экспоненциального обогащения (SELEX) для связывания с их родственной мишенью с высокой аффинностью и специфичностью (48, 49).Библиотеки ДНК содержат модифицированные дезоксиуридиновые основания, которые несут гидрофобные фрагменты в положении 5, что обычно приводит к превосходным связывающим свойствам SOMAmers по сравнению со стандартными аптамерами (50). Во время SOMAscan реагенты SOMAmer образуют стехиометрические комплексы со своими родственными мишенями и в конечном итоге высвобождаются и гибридизуются с массивом (Agilent), что приводит к относительным единицам флуоресценции (RFU) как показание, пропорциональное концентрации соответствующего белка в образце.
Была применена версия SOMAscan, использовавшаяся на момент поступления образцов: 1,129-сплетение в фазе I и 4,000-сплетение в фазе II. SOMAscan — это запатентованный мультиплексный рабочий процесс для обнаружения относительного количества сигналов, представляющих белки, распознаваемые реагентами с медленным изменением аптамера (SOMAmer), который более подробно описан в другом месте (22). Вкратце, реагенты SOMAmer для более чем 4000 белков были созданы с помощью процедуры, известной как систематическая эволюция лигандов путем экспоненциального обогащения (SELEX) для связывания с их родственной мишенью с высокой аффинностью и специфичностью (48, 49).Библиотеки ДНК содержат модифицированные дезоксиуридиновые основания, которые несут гидрофобные фрагменты в положении 5, что обычно приводит к превосходным связывающим свойствам SOMAmers по сравнению со стандартными аптамерами (50). Во время SOMAscan реагенты SOMAmer образуют стехиометрические комплексы со своими родственными мишенями и в конечном итоге высвобождаются и гибридизуются с массивом (Agilent), что приводит к относительным единицам флуоресценции (RFU) как показание, пропорциональное концентрации соответствующего белка в образце. .Анализируемые вещества, измеряемые в SOMAscan с 4 000 плексами, включают секретируемые белки (47%), внеклеточные домены (28%) и внутриклеточные белки (25%) широких биологических групп, таких как рецепторы, киназы, цитокины, протеазы, факторы роста, протеазы. ингибиторы, гормоны и структурные белки.
.Анализируемые вещества, измеряемые в SOMAscan с 4 000 плексами, включают секретируемые белки (47%), внеклеточные домены (28%) и внутриклеточные белки (25%) широких биологических групп, таких как рецепторы, киназы, цитокины, протеазы, факторы роста, протеазы. ингибиторы, гормоны и структурные белки.
Целевая специфичность была подтверждена для подгруппы аналитов с помощью анализов аффинного захвата (раскрывающегося) с использованием иммобилизованных на гранулах SOMAmers с последующей флуоресцентной меткой аналитов и SDS-PAGE анализа, как описано ранее (27).Этот метод был применим только к белкам сыворотки с высоким и средним содержанием из-за его ограниченной чувствительности к белкам с низким содержанием при использовании небольших (<0,5 мл) объемов сыворотки.
Для всех анализов SOMAscan образцы были разделены на аликвоты в матричные пробирки с двумерным штрих-кодом (ThermoScientific), и стандартные разведения сыворотки (0,005%, 1% и 40%) были сделаны для соответствия широкому диапазону концентраций различных белков сыворотки. у нормальных людей и для захвата большого динамического диапазона концентраций белка.Всего было протестировано 1470 образцов, 92,5% из них годны для анализа. Образцы исключались из анализа, если они демонстрировали признаки грубого гемолиза (очевидный красноватый цвет или аномально высокое соотношение гемоглобина и гаптоглобина, измеренное с помощью SOMAscan), как было отмечено для 53 (3,6%) образцов, или если они не прошли анализ из-за технических проблем (малый объем образца, засорение фильтрующей лунки, неполное фоторасщепление или гибридизация), которые потребовали бы чрезмерных средних масштабных коэффициентов нормализации за пределами допустимого диапазона 0.От 4 до 2,5. Повторяющиеся образцы были усреднены и включены для анализа данных, если они не имели противоречивых протеомных или демографических данных ( n = 6). Также были исключены контрольные образцы, которые не были полностью слепыми ( n = 11) или имели недостаточные клинические данные ( n = 8).
у нормальных людей и для захвата большого динамического диапазона концентраций белка.Всего было протестировано 1470 образцов, 92,5% из них годны для анализа. Образцы исключались из анализа, если они демонстрировали признаки грубого гемолиза (очевидный красноватый цвет или аномально высокое соотношение гемоглобина и гаптоглобина, измеренное с помощью SOMAscan), как было отмечено для 53 (3,6%) образцов, или если они не прошли анализ из-за технических проблем (малый объем образца, засорение фильтрующей лунки, неполное фоторасщепление или гибридизация), которые потребовали бы чрезмерных средних масштабных коэффициентов нормализации за пределами допустимого диапазона 0.От 4 до 2,5. Повторяющиеся образцы были усреднены и включены для анализа данных, если они не имели противоречивых протеомных или демографических данных ( n = 6). Также были исключены контрольные образцы, которые не были полностью слепыми ( n = 11) или имели недостаточные клинические данные ( n = 8). Образцы контроля качества и калибратора запускались параллельно и включали нормальных здоровых местных доноров крови. Образцы перемычки использовались для калибровки наборов данных, полученных при различных испытаниях.
Образцы контроля качества и калибратора запускались параллельно и включали нормальных здоровых местных доноров крови. Образцы перемычки использовались для калибровки наборов данных, полученных при различных испытаниях.
Статистический анализ и моделирование. Двусторонний тест t использовался для определения статистических различий в демографических и клинических метаданных между пациентами с ТБ и другими пациентами. Все данные по белкам были преобразованы в логарифмическую форму для стабилизации дисперсии. Нормализация медиана использовалась для корректировки систематической ошибки анализа для конкретного образца, а масштабные коэффициенты варьировались от 0,4 до 2,5. Для всех сравнений использовались непараметрические статистические тесты: тест Колмогорова-Смирнова (KS) для одномерной проверки гипотез и тест Крускала-Уоллиса для межсайтовых сравнений внутри каждой диагностической категории.В то время как статистика KS является величиной без знака, мы сообщаем статистику KS со знаком, где знак плюс или минус указывает направление дифференциального выражения. Коэффициенты ложных открытий Бенджамини и Хохберга использовались для корректировки значений P для множественных сравнений (51). Отбор стабильности с использованием L1-регуляризованной логистической регрессии использовался для выявления стабильных характеристик в присутствии доступных клинических ковариат (52). Кандидаты в биомаркеры были объединены с использованием наивного байесовского классификатора для создания диагностической модели, которая генерирует вероятность того, что пациент болен туберкулезом, с учетом уровней их белковых биомаркеров.Классификация с помощью модели была основана на логарифмических шансах, которые рассчитывались как логарифм шансов = ln (pr TB / pr non-TB ), где pr — вероятность.
Коэффициенты ложных открытий Бенджамини и Хохберга использовались для корректировки значений P для множественных сравнений (51). Отбор стабильности с использованием L1-регуляризованной логистической регрессии использовался для выявления стабильных характеристик в присутствии доступных клинических ковариат (52). Кандидаты в биомаркеры были объединены с использованием наивного байесовского классификатора для создания диагностической модели, которая генерирует вероятность того, что пациент болен туберкулезом, с учетом уровней их белковых биомаркеров.Классификация с помощью модели была основана на логарифмических шансах, которые рассчитывались как логарифм шансов = ln (pr TB / pr non-TB ), где pr — вероятность.
Для каждого этапа исследования 80% обучающих выборок использовались для создания диагностических моделей и перекрестной проверки. Оставшиеся 20% образцов использовались в качестве набора для проверки удержания для расчета оценок производительности модели перед применением моделей к слепым проверочным образцам.
БЛАГОДАРНОСТИ
Мы благодарим FIND за предоставленные образцы из различных географических регионов и за сбор отличных клинических и демографических метаданных.Большое спасибо бывшим и нынешним коллегам в SomaLogic, в том числе Труди Форман, Мередит Госс, Эми Нельсон, Эшли Лоуэлл, Элин Литтон, Деннису Халладей и Джессике Уильямс, за их вклад в создание биохранилища, банка данных и управление проектами. Мы также благодарим Джеффа Картера, Стива Волка и Шери Уилкокс за создание и описание SOMAmer.
SOMAscan и SOMAmer являются зарегистрированными товарными знаками SomaLogic, Inc.
M.A.D.G. является платным консультантом SomaLogic. Все остальные авторы являются сотрудниками или держат опционы SomaLogic.
Мы благодарим Фонд Билла и Мелинды Гейтс за финансирование этого проекта (ОАЭ; OPP1039652). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
- Авторские права © 2017 De Groote et al.

% PDF-1.5
%
103 0 объект
>
endobj
xref
103 112
0000000016 00000 н.
0000003058 00000 н.
0000003208 00000 н.
0000003243 00000 н.
0000003908 00000 н.
0000004059 00000 н.
0000004208 00000 н.
0000004360 00000 н.
0000004643 00000 п.
0000005302 00000 н.
0000005509 00000 н.
0000005764 00000 н.
0000005870 00000 н.
0000006164 00000 п.
0000006201 00000 н.
0000006372 00000 п.
0000006690 00000 н.
0000006802 00000 н.
0000006916 00000 н.
0000006943 00000 н.
0000007353 00000 н.
0000007749 00000 н.
0000008104 00000 п.
0000008131 00000 п.
0000008270 00000 п.
0000008297 00000 н.
0000008617 00000 н.
0000008971 00000 п.
0000010294 00000 п.
0000011210 00000 п.
0000012264 00000 п.
0000012377 00000 п.
0000013386 00000 п.
0000014122 00000 п.
0000014272 00000 п.
0000014446 00000 п.
0000014756 00000 п.
0000015032 00000 п.
0000015406 00000 п.
0000015433 00000 п.
0000016015 00000 п.
0000016106 00000 п.
0000017117 00000 п.
0000017305 00000 п. 0000017400 00000 п.
0000017712 00000 п.
0000018030 00000 п.
0000018261 00000 п.
0000019238 00000 п.
0000019591 00000 п.
0000020577 00000 п.
0000020647 00000 н.
0000020728 00000 п.
0000023644 00000 п.
0000023933 00000 п.
0000024114 00000 п.
0000024404 00000 п.
0000029437 00000 п.
0000029735 00000 п.
0000030663 00000 п.
0000030744 00000 п.
0000030814 00000 п.
0000031016 00000 п.
0000032569 00000 п.
0000035666 00000 п.
0000035747 00000 п.
0000035817 00000 п.
0000036274 00000 п.
0000036564 00000 п.
0000045274 00000 п.
0000045565 00000 п.
0000046115 00000 п.
0000046185 00000 п.
0000046266 00000 п.
0000046437 00000 п.
0000046512 00000 п.
0000046587 00000 п.
0000046707 00000 п.
0000046853 00000 п.
0000049503 00000 п.
0000049637 00000 п.
0000050918 00000 п.
0000051266 00000 п.
0000051303 00000 п.
0000051534 00000 п.
0000051649 00000 п.
0000051834 00000 п.
0000053853 00000 п.
0000054234 00000 п.
0000054673 00000 п.
0000055033 00000 п.
0000061435 00000 п.
0000017400 00000 п.
0000017712 00000 п.
0000018030 00000 п.
0000018261 00000 п.
0000019238 00000 п.
0000019591 00000 п.
0000020577 00000 п.
0000020647 00000 н.
0000020728 00000 п.
0000023644 00000 п.
0000023933 00000 п.
0000024114 00000 п.
0000024404 00000 п.
0000029437 00000 п.
0000029735 00000 п.
0000030663 00000 п.
0000030744 00000 п.
0000030814 00000 п.
0000031016 00000 п.
0000032569 00000 п.
0000035666 00000 п.
0000035747 00000 п.
0000035817 00000 п.
0000036274 00000 п.
0000036564 00000 п.
0000045274 00000 п.
0000045565 00000 п.
0000046115 00000 п.
0000046185 00000 п.
0000046266 00000 п.
0000046437 00000 п.
0000046512 00000 п.
0000046587 00000 п.
0000046707 00000 п.
0000046853 00000 п.
0000049503 00000 п.
0000049637 00000 п.
0000050918 00000 п.
0000051266 00000 п.
0000051303 00000 п.
0000051534 00000 п.
0000051649 00000 п.
0000051834 00000 п.
0000053853 00000 п.
0000054234 00000 п.
0000054673 00000 п.
0000055033 00000 п.
0000061435 00000 п.